离子注入后同源四倍体核雄性不育水稻的育性表达特性黄群策李玉峰摘要以同源四倍体核不育水稻为研究材料,以氮离子束为诱变源完成了离子注入试验。由此认为,氮离子束注入所导致的育性变异在M3代群体内并没有发生进一步的分离。关于同源四倍体核不育水稻的育性稳定性问题前人已经有过公开的文献报道[3]。利用离子注入技术对同源四倍体核不育水稻进行处理后其雄性不育性的变异特征则是值得研究的问题。......
2023-11-28
黄群策 李玉峰
摘要 氮离子注入后在同源四倍体水稻的后代群体内筛选到1株具有双胚苗特征的突变单株.在多个世代的筛选、纯化和鉴定过程中对其多胚苗性状的表现特点进行了观察鉴定.玉米诱发的同源四倍体双胚苗材料在其性状表达特征、性状表达频率和性状表达的条件等方面均显现出一定的特异性.在试验材料的群体内双胚苗发生频率因材料种类不同或发芽温度条件不同而表现出一定的差异性.种子经过去颖壳处理后可以使其双胚苗频率提高32.4%~36.0%.在同源四倍体双胚苗材料中,其苗位特征表现出明显的多样性。
关键词 同源四倍体;双胚苗水稻;形态特征;性状稳定性
从全球水稻遗传改良的现状来看,固定水稻杂种优势效应或简化水稻杂种优势利用的技术程序是育种者所面临的重大难题之一。根据杂交水稻研究的发展趋势,利用生物体的无融合生殖方式有可能解决这一水稻难题[1]。从被子植物无融合生殖研究的近期研究成果来看,通过筛选双胚苗材料有可能寻找到无融合生殖基因或种质资源,在二倍体水平对双胚苗水稻的筛选和研究已经获得了一些有意义的研究结果[2],而在多倍体水平筛选水稻无融合生殖基因或种质资源则为一系法杂交水稻的研究找到了新的研究方向[3]。离子束生物工程作为一门新兴的交叉学科,在农作物遗传改良上的实用性已经被大量的试验结果所证实[4,5]。经过20多年的探索之后,在离子束生物工程的研究领域内研究者面临的一项重要任务就是对具有特异性的突变体材料的性状表达规律进行更深入地研究.本项研究以氮离子束为诱变源,对同源四倍体水稻进行注入处理后在其当代群体内获得了同源四倍体双胚苗突变体.以此为基础,在对突变体后代进行筛选和纯化的过程中对其双胚苗的形态特征及其性状表达频率进行了研究,旨在更好地认识在同源四倍体水平双胚苗性状的表达特点,进而为更深入地开展同源四倍体水稻的研究和促进离子束生物工程的发展积累更多的参考资料。
1 材料与方法
1.1 氮离子注入试验
在氮离子注入试验中以两种不同染色体组倍性的水稻[即同源四倍体水稻品系99-01(4)和相应的二倍体水稻品种99-01(2)]作为试验材料。前者是以后者为亲本材料通过种芽诱导法所筛选到的主要农艺性状已经达到稳定状态的同源四倍体水稻品系[6]。在进行离子注入试验之前,每份试验材料均进行了纯化试验,以便证实其遗传上的稳定性。

图1 具有双胚功特征的同源四倍体水稻
在试验中以能量为25 keV的氮离子束为诱变源,离子注入剂量为4×1016N+/cm2,在真空靶室内对试验材料的种胚进行离子注入处理。在试验中设两个对照,即不进入真空靶室内的空白对照(CK1)和在真空靶室内不经过离子注入处理的对照(CK2)。在离子注入处理中所采用的仪器设备为由中国科学院等离子体物理研究所自制的LCD-1000多功能离子注入机。在进行离子注入试验中首先将每份水稻种子1 000粒去壳,将其插入并固定在盛有花泥的培养皿中。种子的胚部露在花泥的外面(即种胚朝上露出),以便能直接受到氮离子束的照射。
在静真空度和工作真空度均为10-6Pa的条件下进行离子辐照处理。随后,按照常规水稻的浸种催芽方法在30℃的条件下进行浸种和催芽,再将已经成苗的实验材料寄栽到实验盆内。当实验材料具有5~6片真叶时,再按照30cm×25cm行株距将其移栽至大田,让其在自然条件下生长发育。
1.2 对双胚苗突变体材料的筛选
在经过离子注入处理的试验材料进入成熟期时,分别从中筛选出一些外观农艺性状比较特殊的变异单株,在隔离条件下让其自交繁殖并以试验材料的单株为单位进行留种,由此形成第二代群体(M2)。在对M2材料的发芽状态进行鉴定时发现了1株具有双胚苗特征(即1粒种子同时萌发出两株种苗)的种子(图1)。将具有双胚苗特征的材料种植在试验盆内,对两株种苗分别进行标记(其中包括由每一主茎所产生的分裂植株)。由每一主茎植株及其所产生的分裂植株组成一个遗传单位进行留种,由此在成熟期形成两份M3后代材料(即M3-1和M3-2)。在两份M3后代种子进行催芽期间开始对表现双胚苗特征的材料进行鉴定和统计,由此确定在两个后代群体内双胚苗性状在形态学上的特异性及其表现频率。在对突变体后代进行筛选的过程中按照试验材料的形态学特征进行单株归类和留种。在田间条件下对试验材料的主要农艺性状进行筛选,以单株为单位留种,由此形成M4后代材料。从M4群体开始直至M6群体,以主要农艺性状的表现状况为鉴定指标进行单株筛选和后代留种,并对其双胚苗的特异性及其表现频率进行鉴定和统计。
1.3 对双胚苗形态特征的观察鉴定
从M3群体开始到M6群体,以单株为单位对其双胚苗发生频率进行了鉴定,在观察鉴定试验中,首先按照试验设计将各份试验材料的种子放入恒温为30℃的清水中浸泡2 d。随后,将其放入铺有湿润滤纸的培养皿中催芽和培养,10 d后分别统计其群体内双胚苗、三胚苗和四胚苗的数目,由此计算多胚苗的发生频率(双胚苗频率= ×100%;三胚苗频率=
×100%;三胚苗频率= ×100%;四胚苗频率=
×100%;四胚苗频率= ×100%;多胚苗频率=双胚苗频率+三胚苗频率+四胚苗频率)。
×100%;多胚苗频率=双胚苗频率+三胚苗频率+四胚苗频率)。
对于同源四倍体水稻群体内所出现的双胚苗突变材料,按照单株筛选法对其进行3个世代的筛选和纯化,待其主要农艺性状稳定后按照试验设计对其双胚苗的形态特征及其性状稳定性进行了研究。在试验中以同源四倍体双胚苗株系ASDOR05-01和ASDOR05-02同源四倍体水稻99-01(4)和相应的二倍体水稻99-01(2)为研究材料。试验设计包括4次重复,在每一次重复中每份试验材料的4000粒种子分为留壳和去壳两种处理,选用4种发芽温度(即20℃、25℃、30℃和35℃)对试验材料的特征特性进行观察鉴定,统计其双胚苗的发生频率。在对试验材料的双胚苗发生频率进行鉴定的同时,对双胚苗的形态特征进行观察鉴定。
2 结果与分析
2.1 氮离子注入后在当代群体和M2代群体内的生物学效应
在CK1和CK2的当代群体内没有发现具有明显变异特征的突变体,群体的主要农艺性状表现为整齐一致.在经过氮离子束注入后的当代群体内出现了一些具有生育特性和形态学特异性的突变体,其突变体的类型和变异频率因材料种类不同而异,性状的变异并没有一定的规律性,而是表现出明显的随机性。
在同源四倍体水稻品系99-01(4)的后代群体内所出现的突变频率(9.6%)明显超过在二倍体水稻品种99-01(2)的后代群体内所出现的突变频率(1.2%)。突变体的生育特性主要涉及生育期的长短,其中包括早熟类型、迟熟类型和中间类型。突变体的形态学特异性主要涉及植株高度、剑叶长短和宽窄、穗长、穗粒数和叶色等。在试验中,对于各种突变体材料按照不同材料类型单株收种。
在二倍体水稻品种99-01(2)的M2代群体内存在着一些表现为形态学变异的突变单株,但没有发现具有双胚苗特征的突变株。在经过氮离子束注入处理的同源四倍体水稻99-01(4)的M2代群体内除了存在着比较多的突变单株之外,特别发现了一株具有双胚苗特征的突变株[当时将其命名为99-01(4)-双]。根据当时的观察鉴定结果,该突变株所形成的两株苗表现为均势双苗,即形成的两株苗在生长势上大小相当,没有表现出强弱之分。以该双胚苗突变株的不定根根尖为材料进行染色体鉴定的结果表明,在这两株苗的细胞内所携带的染色体数目均为48条。将双胚苗植株中的两个单株分别进行标记后,在成熟期获得了由两个单株分别产生的M3群体。
2.2 在M3群体至M5群体内双胚苗性状的遗传动态
在试验中通过对M2群体内的双胚苗材料进行标记和筛选后获得两份M3群体。在M3群体内各个单株在主要农艺性状上仍然表现出明显的分离现象。与其原始亲本相比,99-01(4)-双的主要农艺性状有其特异性(表1)。发芽试验的统计结果表明,在M3群体内除了存在着双胚苗单株之外,还出现了三胚苗单株和四胚苗单株,其多胚苗频率为12.50%。在全部的多胚苗材料中,双胚苗单株、三胚苗单株和四胚苗单株出现的相对频率分别为81.95%,9.72%和8.33%。然而,在随后的世代中在群体内没有发现三胚苗材料和四胚苗材料(表2)。
表1 99-01(4)-双的主要农艺性状(www.chuimin.cn)

表2 在M3~M5代群体内IR36-双的多胚功频率

在M4群体内,在主要农艺性状的表现上,99-01(4)-双与其对照单株仍然存在着一定的差异,与上一个世代的双胚苗单株也存在着明显的差异,这说明其主要农艺性状处于进一步的分离状态。对双胚苗单株的发芽试验结果表明,在2 340粒种子中多胚苗总数为297株(12.69%),其中包括的双胚苗、三胚苗和四胚苗的数目分别是246株(10.51%)、31株(1.32%)和20株(0.86%)。
在M5群体内,99-01(4)-双株系的主要农艺性状趋于稳定。在试验中以M4群体内的均势双胚苗单株的自交后代组成M5群体进行发芽试验。统计结果表明,在3 000粒种子中没有发现三胚苗个体或四胚苗个体,具有双胚苗特征的种子数目为273粒,其双胚苗频率为9.10%。
2.3 在M6稳定群体内双胚苗性状表达的特异性
在M6群体内各个株系之间在主要农艺性状上存在着一定的差异,但在株系内各个单株之间其主要农艺性状相对稳定.在试验中以具有双胚苗特征的两个整齐株系(即ASDOR05-01株系和ASDOR05-02株系)为研究材料,对其双胚苗的发生频率进行了观察鉴定和统计分析.研究结果表明,作为对照的同源四倍体水稻99-01(4)和相应的二倍体水稻99-01(2)在4种温度条件下进行发芽试验,在群体内均没有发现具有双胚苗特征的个体,这说明它们不具有双胚苗特性。在两份双胚苗株系中均出现了双胚苗个体,但在群体内双胚苗的发生频率因材料种类不同而异,或因同一种材料在不同的发芽温度条件下其双胚苗的频率也表现出一定的差异(表3)。从双苗性状发生频率的总体平均值来看,ASDOR05-01的双胚苗频率(4.3%)要高于ASDOR05-02的双胚苗频率(1.3%)。在发芽温度为30的条件下,ASDOR05-01和ASDOR05-02的双胚苗频率均为最高值,分别为6.8%和2.5%。
表3 试验材料的发芽率和双胚苗率 %
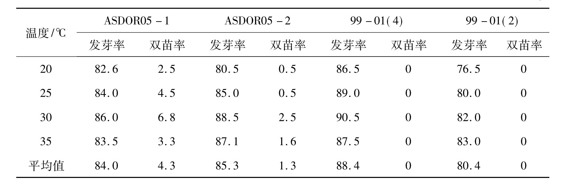
根据以前的研究结果,水稻种子颖壳的有无对其双胚苗频率和发芽率有一定的影响[2]。在本试验的研究中,试验材料在25℃条件下浸种,在30℃条件下进行催芽,对于试验材料颖壳的有无所导致的效应进行了研究。研究结果表明,对于所有的试验材料而言,种子去壳有利于提高其发芽率,发芽率提高的幅度在30%以上。在两份对照材料[99-01(4)和99-01(2)]中没有发现双胚苗个体,而在ASDOR05-01和ASDOR05-02群体内均出现了双胚苗个体。在ASDOR05-01群体内,种子去颖壳处理后使其双胚苗频率提高至9.0%,这比同样条件下种子不去颖壳的处理结果(6.8%)提高了32.4%。在ASDOR05-02群体内,种子去颖壳处理后使其双胚苗频率提高至3.4%,这比同样条件下种子不去颖壳的处理结果(2.5%)提高了36.0%。由此可见,在水稻双胚苗频率的鉴定试验中种子颖壳的有无在一定程度上影响着其双胚苗性状的表达频率。
根据对双苗材料的苗位进行观察的结果表明,其双胚苗的苗位存在着两种类型,即非完全双胚苗和完全双胚苗。在非完全双胚苗中主要有单胚轴单胚根双胚苗和单胚根异胚轴双胚苗两种类型。在完全双胚苗中有正常双胚苗(即双苗的生长势比较均匀,没有明显的大小苗之分)和异常双胚苗(即双苗的生长势很不均匀,存在着明显的大小苗之分)。由此可见,在同源四倍体双胚苗材料中,其苗位特征表现出明显的多样性和差异性。
3 讨论
在确保水稻产量潜力的前提下如何简化利用水稻杂种优势的技术程序,即固定水稻的杂种优势是水稻遗传改良者目前所面对的难题之一。前人在禾本科植物遗传改良的研究中发现,固定禾本科植物杂种优势的途径有多条,其中最好的途径就是有效地利用无融合生殖种质资源,进而建立挖掘其无融合生殖潜力的育种技术程序[1,3]。从当前的研究趋势来看,各国研究者已经将选育具有无融合生殖特性的农作物品种作为掀起又一次绿色革命浪潮的突破口而对其给予高度重视,正加紧在高梁、玉米、小麦、珍珠粟和水稻等农作物中开展无融合生殖的研究。我国水稻无融合生殖的研究主要集中在筛选和鉴定多胚苗材料,对于多胚苗水稻的形态学、胚胎学和遗传学的研究已经获得了一些具有学术价值的研究资料[6]。然而,前人关于多胚苗水稻的研究主要在二倍体水平展开探索,而关于同源四倍体多胚苗水稻的研究目前尚未见到公开的文献报道[3]。在禾本科植物中具有无融合生殖特性的大多数物种,在染色体组水平上表现出多倍化的特征,由此促使研究者将寻找水稻无融合生殖种质的研究重点由二倍体水稻转向多倍体水稻。
利用离子束注入技术对同源四倍体水稻进行遗传改良的实用性已经被大量的研究结果所证实[7]。通过离子束注入之后在其后代群体内筛选到具有双胚苗特性的突变材料,对其双胚苗形态特征及其性状稳定性值得进一步研究。研究结果表明,通过离子束注入处理后在同源四倍体水稻群体内所获得的具有双胚苗特性的突变材料,在其自交后代株系内仍然按照一定的频率保持着双胚苗特性,其变异特征属于遗传性变异。随着自交世代的推进,在其主要农艺性状趋于稳定的同时,其双胚苗特性的表达频率发生了一定的变化。在高世代的两份多胚苗株系中均出现了双胚苗个体,但群体内双胚苗发生频率因材料种类不同或发芽温度条件不同而表现出一定的差异性。从双苗性状发生频率的总体平均值来看,ASDOR05-01的双胚苗频率(4.3%)要高于ASDOR05-02的双胚苗频率(1.3%)。在发芽温度为30℃的条件下,ASDOR05-01和ASDOR05-02的双胚苗频率均为最高,分别为6.8%和2.5%。在双胚苗频率的鉴定试验中种子颖壳的有无在一定程度上影响着其双胚苗性状的表达频率。在ASDOR05-01群体内,种子去颖壳处理后使其双胚苗频率提高至9.0%,这比同样条件下种子不去颖壳的处理结果(6.8%)提高了32.4%。在ASDOR05-02群体内,种子去颖壳处理后使其双胚苗频率提高至3.4%,这比在同样条件下种子不去颖壳的处理结果(2.5%)提高了36.0%。在同源四倍体双胚苗材料中,其苗位特征表现出明显的多样性。由此可见,利用离子注入技术对同源四倍体水稻进行改良则可以获得一些具有遗传性变异特点的新种质,同源四倍体双胚苗材料在其性状表达特征、性状表达频率和性状表达的条件等方面均显现出一定的特异性。然而,关于ASDOR05-01和ASDOR05-02的特征特性还有许多问题值得进一步深入研究。
[1]袁隆平.杂交水稻超高产育种[J].杂交水稻,2001,16(1):1-3.
[2]郭学兴.中国水稻无融合生殖研究进展[M].成都:四川科学技术出版社,1991.
[3]黄群策.被子植物的无融合生殖[M].福州:福建科学技术出版社,2000.
[4]余增亮.离子束生物技术引论[M].合肥:安徽科学技术出版社,1998.
[5]黄群策,李玉峰.离子束生物技术在水稻育种中的应用前景[J].杂交水稻,2002,17(5):53-56.
[6]黄群策,孙敬三,朱生伟.种芽诱导获得同源四倍体水稻技术的研究[J].中国农学通报,1997,3(6):21-23.
[7]黄群策,秦广雍.离子束生物技术改良同源四倍体水稻的设想[J].郑州大学学报,2003,35(4):31-36.
【注释】
(1)该文曾在《湖南农业大学》 [2007,33(2):46~50]刊出
有关水稻染色体组多倍化研究的文章

离子注入后同源四倍体核雄性不育水稻的育性表达特性黄群策李玉峰摘要以同源四倍体核不育水稻为研究材料,以氮离子束为诱变源完成了离子注入试验。由此认为,氮离子束注入所导致的育性变异在M3代群体内并没有发生进一步的分离。关于同源四倍体核不育水稻的育性稳定性问题前人已经有过公开的文献报道[3]。利用离子注入技术对同源四倍体核不育水稻进行处理后其雄性不育性的变异特征则是值得研究的问题。......
2023-11-28

同源四倍体双胚苗水稻雌配子体的发育特征黄群策代西梅贾宏汝摘要利用激光扫描共聚焦显微技术对3份水稻材料的雌配子或雌配子体的发育特征进行了观察鉴定。2结果与分析2.1试验材料正常雌配子体的发育特征根据前人的研究结果,水稻雌配子体的正常发育过程有其特点。......
2023-11-28
从低能离子束生物工程的发展趋势来看,对于突变体材料所表现出的后效性值得关注,因而也对于同源四倍体双胚苗水稻的生殖发育特性值得进行深入研究。关于二倍体水稻的双胚苗的形态特征及其生殖发育特性,前人已经进行过一些研究[4-6],而关于同源四倍体双胚苗水稻的雄配子体发育特性尚未见到公开的文献报道。具体见上篇“同源四倍体双胚苗水稻的生殖发育特征研究”。......
2023-11-28

同源四倍体双胚苗水稻的生殖发育特征研究黄群策黄雅琴代西梅摘要利用激光共聚焦显微扫描技术对同源四倍体双胚苗水稻品系ASDOR05-01的雄配子体和雌配子体的发育特征进行了观察鉴定。研究结果表明,在每1份试验材料中均存在着正常的雌雄配子体和异常的雌雄配子体,而同源四倍体双胚苗水稻品系ASDOR05-01表现出更弱的有性生殖能力。......
2023-11-28

研究结果表明,同源四倍体水稻和二倍体水稻在发生受精作用的时间上没有存在着很大的差异。同源四倍体水稻在生殖特性上表现出一定的特异性,其研究价值和潜在的利用价值有待于进一步探索[3]。本项研究利用激光扫描共聚焦显微技术对同源四倍体双胚苗水稻的受精过程、胚胎发育过程和双胚来源进行了观察鉴定,旨在研究其生殖发育特性并寻找双胚苗发生的胚胎学证据。......
2023-11-28
氮离子注入对同源四倍体水稻成熟胚组织培养体系的影响李玉峰黄群策梁运章摘要选择3份同源四倍体水稻为材料,研究了氮离子注入对其成熟胚诱导愈伤组织和分化成苗的影响。本文通过对四倍体水稻成熟胚进行氮离子注入处理,探讨了离子注入对四倍体水稻成熟胚组织培养体系影响的规律,从而为以后的应用研究做好理论铺垫和方法准备。并且在不同的注入剂量范围,愈伤组织增重倍数增加的趋势有所不同。......
2023-11-28

离子注入后同源四倍体多胚苗突变水稻的筛选及其遗传稳定性代西梅黄群策胡秀明摘要以低能氮离子束为诱变源,对同源四倍体水稻品系“IR36-4X”进行离子注入,在其第2代群体内筛选得到了1株具有多胚苗性状特征的突变株。并对该突变株系的苗位、主要农艺性状和多胚苗遗传稳定性进行了跟踪调查。......
2023-11-28

黄群策等[6]以低能氮离子束为诱变源对同源四倍体水稻进行了遗传改良,在其后代群体内筛选到具有双胚苗特征的新材料为了对同源四倍体双胚苗突变水稻进行有效的遗传改良,郑州大学离子束生物工程河南省重点实验室的研究人员利用无性系诱导技术对其脱分化和再分化特性进行了初步研究,试图为在细胞工程水平研究同源四倍体水稻双胚苗材料的细胞学特性提供技术资料。......
2023-11-28
相关推荐