双胚苗水稻种子的扫描电镜观察黄雅琴黄群策燕晓阳摘要以同源四倍体双胚苗水稻品系及其衍生后代为研究材料,利用扫描电镜技术和X射线衍射技术对稻米的垩白性状淀粉粒的形态特征和淀粉的晶体特性进行了初步研究。关键词双胚苗水稻;同源四倍体;米质;淀粉粒;扫描电镜; X射线衍射仪水稻是世界上最主要的粮食作物,选育高产、优质、多抗统一的水稻新品种是21世纪的育种目标[1]。......
2023-11-28
代西梅 黄群策 李国平
摘要 利用激光扫描共聚焦显微镜术对水稻品种IR36二倍体及其同源四倍体的胚囊形成和发育过程进行了系统的研究。在IR36二倍体胚囊的发育过程中,观察到了几种前人未曾报道的异常胚囊类型,即三核胚囊、六核胚囊、九核胚囊及一些退化胚囊,其形成机制有待进一步研究。观察结果可为正确认识水稻胚囊发育形成过程提供参考资料。
关键词 水稻;胚囊;激光扫描共聚焦显微镜;发育
利用激光扫描共聚焦显微镜可以对样品材料进行连续扫描,通过电脑处理从而获得其三维立体结构,使我们对其样品内各细胞的空间分布有更清晰的认识。激光扫描共聚焦显微镜的应用为水稻胚囊结构、形成及发育过程等方面的研究提供了全新的视野[1,2]。笔者利用激光扫描共聚焦显微镜术对水稻胚囊的形成及发育过程进行了较为系统的观察。在对水稻IR36二倍体及其同源四倍体胚囊发育的观察过程中,发现了一些前人未曾报到过的特殊现象,这对我们正确认识水稻胚囊的形成发育过程将会有一定的参考价值。
1 材料和方法
试验材料为水稻IR36的二倍体材料及同源四倍体材料(IR36同源四倍体胚囊发育过程将作后续报道)。2004年将试验材料种植于新乡市农业科学院水稻研究所的试验田内,常规管理。在水稻胚囊发育的各个时期,取穗中部小穗固定于FAA固定液中,每个时期约取小穗100个。观察前用70%的酒精冲洗一下,接着在解剖镜下分离出子房,保存于70%酒精中备用。染色前,各个时期的水稻子房经50%、30%、15%乙醇及蒸馏水冲洗,每次20min。然后在60℃水浴中用1mol/L的HCl水解10min左右。1%曙红Y衬染8h左右,加入蒸馏水冲洗,换液数次至无浮色。柠檬酸-磷酸氢二钠缓冲液(0.1mol/L,pH = 5.0)预处理(8h左右),20 g/mLH33342(Hoechest s tain)染色24h(25℃黑暗)。用蒸馏水冲洗2~3次,乙醇梯度(15%、30%、50%、70%、85%、95%)脱水,每级20min,随后将子房放入无水乙醇中脱水2次,每次的脱水时间为2h,接着转入无水乙醇中过夜。第2天将已经脱水的材料放入由无水乙醇和水杨酸甲酯(1∶1)组成的混合液中过渡1h,再用水杨酸甲酯透明处理3次。在前2次透明时,每次2h,最后一次透明的时间为15h。所有的试验材料经过水杨酸甲酯透明处理后可以在水杨酸甲酯中保存备用。观察前,用镊子轻轻将处理好的子房夹出,置于凹玻片上,用丁香油封片(注意加盖玻片时,不要挤压,防止子房变形或破裂)。将制备好的载玻片倒置于Leica SP2激光共聚焦扫描显微镜上,用488nm波长的激光激发,扫描获得清晰的胚囊各个时期的图片。
2 结果与分析
水稻胚囊属于蓼型胚囊,胚囊的形成发育过程经过孢原细胞发育成大孢子母细胞,大孢子母细胞经过2次减数分裂形成四分体(图1-A),四分体中珠孔端的3个细胞逐渐退化(图1-B,图1-C),合点端的细胞成为功能大孢子,功能大孢子进一步发育成为单核胚囊(图1-D),然后单核胚囊经过3次有丝分裂,经二核胚囊(图1-E),四核胚囊(图1-F),最后发育形成八核胚囊的结构(图1-G,图1-H)。图1-G和图1-H为同一个胚囊的2个不同扫描切面。通过激光共聚焦扫描显微镜的连续扫描功能,可以对胚囊内各个核的空间分布有更进一步的立体认识。笔者通过对大量的水稻IR36二倍体胚囊进行整体观察,发现一些前人未曾报道或与前人报道结果不一致的现象。大孢子母细胞经过减数分裂形成四分体,随后,四分体中靠近珠孔端的3个细胞依次退化。本文提供的激光扫描共聚焦显微镜图像首次清晰地显示了四分体中珠孔端3个细胞依次退化的过程,其解体后的细胞显示出强烈荧光(图1-B,图1-C,图1-D)。Webb等[3]研究认为这可能是由于胼胝质在退化细胞的细胞壁积累所致。
刘向东等[4]研究认为,大孢子母细胞减数分裂末期2次胞质分裂都是不均等的,本人在研究中也观察到了类似现象。但刘向东等[4]指出,四分体中,从珠孔端到合点端,细胞依次变大,即合点端的细胞最大,珠孔端的细胞最小。笔者的观察结果(图1-A)不能支持刘向东的描述。
应用激光扫描共聚焦显微镜证实了在胚囊形成发育过程中出现了三核胚囊、六核胚囊、九核胚囊及退化胚囊。刘向东等[4]曾明确指出,第2,3次分裂时,胚囊内各核同步分裂,因而经历二核胚囊、四核胚囊至八核胚囊,完全不存在三、五和六核胚囊。本研究过程中清晰观察到了三核胚囊、六核胚囊及九核胚囊的存在,三核胚囊中没有大液泡的形成,核也没有移向胚囊的两端,3个核均位于胚囊的中部(图1-I)。六核胚囊的6个核中,3个核位于珠孔端,另外3个核位于合点端。珠孔端及合点端各有1个核有向胚囊中央移动的倾向(图1-J)。九核胚囊中5个核位于珠孔端,4个核位于合点端(图1-K,L)。图1-K和图1-L为同一个胚囊的2个连续激光扫描切面,图K显示合点端的4个核,图1-L显示珠孔端的5个核。另外在观察过程中还发现了一些退化胚囊,胚囊中没有细胞结构,只显示细胞解体后的强烈荧光(图片未提供),这与四分体中珠孔端3个细胞退化后的现象有些相像。
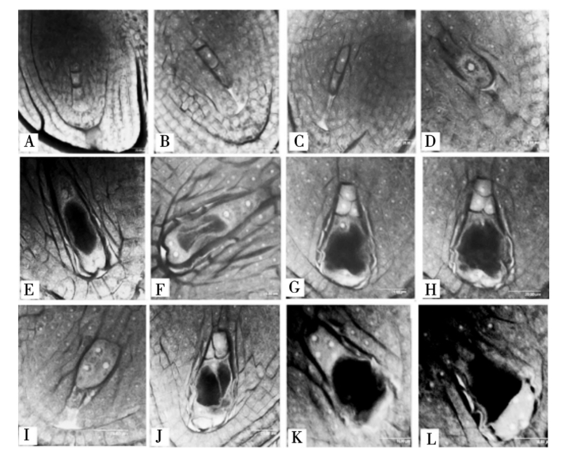
图1
A.四分体; B.珠孔端的2个细胞已经退化; C.珠孔端的第3个细胞开始退化; D.单核胚囊; E.二核胚囊; F.四核胚囊; G,H.早期八胚囊(同一胚囊的不同切面); I.三核胚囊; J.六核胚囊; K,L.九核胚囊的2个不同切面,K.表示合点端的4个核,L.表示孔端的5个核
3 讨论
戴伦焰等[5]在对水稻胚囊的形成与发育的研究中明确了水稻胚囊的形成发育是蓼型即单孢子、八核、二等级等,并描述了水稻从孢原细胞到成熟胚囊的发育过程。随后,Hoshikawa[6 ]也对水稻胚囊的形成过程进行了较为系统的研究,并将其形成过程分为12个时期。刘向东等[4]在前人研究的基础上结合自己大量的研究事实将水稻胚囊的形成过程分为8个时期,并对每个时期的主要形态特征作了详细描述。这对我们认识水稻胚囊的形成发育过程有一定的指导意义。但是,根据以往学者们对水稻胚囊的认识,大都认为水稻单核胚囊形成后,经过第1次有丝分裂,形成二核胚囊。二核胚囊再经过同步分裂,形成四核胚囊。然后,再次经过同步分裂形成八核胚囊。由于第2次和第3次有丝分裂均是同步分裂,所以在水稻胚囊内绝对不会有三核胚囊、五核胚囊和六核胚囊。然而,作者在研究过程中,确实观察到了清晰的三核胚囊、六核胚囊及九核胚囊的存在。这说明有必要对水稻胚囊发育过程进行更深入的研究。
刘向东等[7]在研究多胚水稻品系APIV胚囊的形成与发育的过程中,在其变异型胚囊5-2-1型、6-2-0及5-3-0的发育过程中都曾发现三核胚囊的存在。5-2-1型胚囊的3个核一个位于合点端,另外2个位于珠孔端;而6-2-0及5-3-0胚囊的3个核均位于珠孔端。认为变异胚囊的形成是核的异常行为及胚囊核的不同步分裂造成的。并指出变异型胚囊发育过程中核异常行为受着微管骨架组织变化的影响,显示微管骨架组织在胚囊核行为中起着一定的作用[8]。笔者在IR36胚囊发育过程中观察到的三核胚囊类似于多胚水稻品系APIV变异胚囊6-2-0及5-3-0的三核胚囊,3个核均位于胚囊的中部而没有移向两端,在胚囊中也没有发现大液泡的形成。(www.chuimin.cn)
Haga[9]及Malik[10 ]研究均指出,当植物由原产地迁移到另一地区时,由于环境条件的改变,会引起其分裂发生异常。吴素萱等[11]研究也指出,无论是生物的、生理的、遗传的以及各种不正常的环境因素都可以引起细胞内部的变化,这可能是上述因素改变了细胞内物质的正常代谢过程,因而导致细胞分裂时染色体的不相适应或不相协调。笔者认为,上述因素对大孢子的形成以及胚囊的发育也可能有一定的影响。
三核胚囊的形成可能由于环境条件的改变,影响了胚囊细胞内各种代谢过程,引起胚囊内微管骨架组织的变化,在形成二核胚囊后2个核不能正常的移向胚囊的两端,而后是2个核的不同步分裂,其中1个核先分裂,因而在胚囊内就出现了3个核的现象。六核胚囊的形成也可能是由于形成四核胚囊后核的不同步分裂造成的。九核胚囊的形成则可能是一种变异现象。至于出现本文所描述的特异胚囊类型的原因,还有待于进一步的研究。
[1]任宏,刘永胜,孙敬三.用共聚焦扫描显微镜检术观察水稻胚囊发育[J].植物学报,1998,40(9):786-789.
[2]张华华,冯九焕,卢永根,等.利用激光扫描共聚焦显微镜观察同源四倍体水稻胚囊的形成与发育[J].电子显微学报,2003,22(5):380-384.
[3]Webbm C,GunningB E S.Embryo sac development inArabidopsis thaliana.Megasporogenesis,including themicrotubularcytoskeleton[J].Sex Plant Reprod,1990,(3):244-256.
[4]刘向东,徐雪宾,卢永根,等.水稻胚囊形成过程与分期[J].中国水稻科学,1997,11(3):141-150.
[5]戴伦焰,曾子申.水稻胚囊的形成与发育[J].武汉大学学报(自然科学版),1964,2(1):97-110.
[6]Kiyochikahoshi kawa.The growing rice plant(an anatomicalmonograph)[M].Tokyo: Tokyo Nobunkyo,1989: 220-223.
[7]刘向东,卢永根,徐雪宾,等.多胚水稻品系不同类型胚囊的形成与发育[J].植物学报,1996,38(10):767-771.
[8]LIU Xiangdong,LU Yonggen,ZHUhong-Liang,et.al.Abnormal behavior of nuclei andmicrotubule(MT)organizational changes during embryosac development in the polyeggmutantAPIVof Rice[J].ActaBotanicaSinica,2004,46(7):829-838.
[9]Haga T.Chromosome complement of kinugasa japonica withspecial reference to its origin and behavior[J].Cytologia,1937,8(1):137-141.
[10]Malik C P,Tandon S L.Cytological behaviour of Asparagurcurillusham.Inrelation to different environments[J].TheIndian Jhort,1959,16(1):39-41.
[11]吴素萱,蔡起贵.环境条件对于水稻减数分裂的影响[J].植物学报,1962,10(1):1-10.
【注释】
(1)该文曾在《华北农学报》 [2006,21(6):26~29]刊出
有关水稻染色体组多倍化研究的文章

双胚苗水稻种子的扫描电镜观察黄雅琴黄群策燕晓阳摘要以同源四倍体双胚苗水稻品系及其衍生后代为研究材料,利用扫描电镜技术和X射线衍射技术对稻米的垩白性状淀粉粒的形态特征和淀粉的晶体特性进行了初步研究。关键词双胚苗水稻;同源四倍体;米质;淀粉粒;扫描电镜; X射线衍射仪水稻是世界上最主要的粮食作物,选育高产、优质、多抗统一的水稻新品种是21世纪的育种目标[1]。......
2023-11-28

FLC和CO对SOC1的调控是通过SOC1的启动子区域进行转录调控。FT也是传递开花信号的一个主要基因,受CO的正调控和FLC的负调控,促进开花关键基因AP1的表达。LFY同样也是长日和赤霉素信号的产物,这些信号也是通过LFY启动子区域进行转录水平的调控。AP1和LFY是最终决定花分生组织特性的关键基因,它们的表达,形成了花原基,称为花分生组织,决定了花结构形成的起始。Blazquez将已经获得的相关基因的关系绘制了一张网络图综合说明花发育的基因决定过程。......
2023-11-20

精液是碱性的,女性生殖器内部则为酸性。至第八周末,所有的器官已初具原形。第八周初,胎头占整个胎儿全长的二分之一,以后生长加快。至第十二周末,已显示成熟胎儿男女外阴的形态。皮肤出现胎毛,心率是成人的两倍。视频中儿子给去世的妈妈做了一顿妈妈盼望已久的午餐,却只能是送往天堂的午餐。看完《天堂的午餐》,我想说……......
2024-01-28

受精启动果实的发育。授粉以后,果实的发育也与种子发育密切相关。在李子发育过程中,生长素在果实生长中起重要作用,而乙烯在果实成熟的起始和速度方面起重要作用。赤霉素Gibberellin在受精后角果的发育过程中促进角果的延长,缺乏赤霉素时抑制生长。GAI编码转录调控辅助因子,17个氨基酸的去除造成对赤霉素不敏感的突变gai。gal-5gai突变的表型与未授粉但用生长素处理的表型相同,说明GA缺乏时,生长素的作用占主导地位。......
2023-11-20

由此认为,同源四倍体水稻的有性生殖能力已经明显变弱。关于二倍体水稻的受精过程的细胞学观察,前人已有报道[5-7]。但是,关于同源四倍体水稻的双受精过程,目前国内外尚未见到公开的文献报道。对二倍体水稻和同源四倍体水稻的双受精过程进行比较研究将有助于了解其生殖特性的差异,这对于从同源四倍体水稻中鉴定和筛选潜在的无融合生殖基因具有重大的现实意义。......
2023-11-28

开花植物中,胚囊的种类有四孢子起源、二孢子起源、单孢子起源,即使是单孢子起源,核在胚囊内迁移的数目和位置决定了中心细胞的核数目,从而决定受精后胚乳的倍性。图6.9介绍了不同进化水平的植物胚囊结构、中心细胞核数目和双受精后胚乳的倍性。下面以双子叶植物拟南芥和单子叶植物玉米为代表介绍胚乳的形成过程和机理。受精时一个来自花粉管的单倍体雄核进入中心细胞,三个核融合形成三倍体原胚乳核。......
2023-11-20

研究结果表明,同源四倍体水稻和二倍体水稻在发生受精作用的时间上没有存在着很大的差异。同源四倍体水稻在生殖特性上表现出一定的特异性,其研究价值和潜在的利用价值有待于进一步探索[3]。本项研究利用激光扫描共聚焦显微技术对同源四倍体双胚苗水稻的受精过程、胚胎发育过程和双胚来源进行了观察鉴定,旨在研究其生殖发育特性并寻找双胚苗发生的胚胎学证据。......
2023-11-28

为了保存这些资料,系领导决定让我留守宜山,负责观察记载各品种特性和田间试验结果等工作,并派农工老章协助田间除草、施肥、收获等工作。学校于5月下旬派一辆柴油大卡车到宜山,我和农工将种子箱和学校留存在农场的少量物品,装满一车,并带着县农场退还的租金2000元支票离别宜山,直开遵义。二天后才到达遵义本部,完成了留守宜山的任务。......
2023-10-07
相关推荐