同源四倍体双胚苗水稻雌配子体的发育特征黄群策代西梅贾宏汝摘要利用激光扫描共聚焦显微技术对3份水稻材料的雌配子或雌配子体的发育特征进行了观察鉴定。2结果与分析2.1试验材料正常雌配子体的发育特征根据前人的研究结果,水稻雌配子体的正常发育过程有其特点。......
2023-11-28
同源四倍体双胚苗水稻的生殖发育特征研究
黄群策 黄雅琴 代西梅
摘要 利用激光共聚焦显微扫描技术对同源四倍体双胚苗水稻品系ASDOR05-01的雄配子体和雌配子体的发育特征进行了观察鉴定。研究结果表明,在每1份试验材料中均存在着正常的雌雄配子体和异常的雌雄配子体,而同源四倍体双胚苗水稻品系ASDOR05-01表现出更弱的有性生殖能力。同源四倍体双胚苗水稻品系ASDOR05-01在小孢子母细胞进行减数分裂时,出现了5种不正常的分裂现象,即部分染色体不能正常配对、在分裂中期形成了三极纺锤体、形成异常的二分体、在第二次减数分裂中期出现了分裂不同步现象、形成异常的四分体。在ASDOR05-01植株的雌配子体发育过程中,其特异性主要包括11种类型。同源四倍体水稻和二倍体水稻在发生受精作用的时间上没有存在着很大的差异。双胚苗水稻的受精作用和胚胎发育表现出一定的特点,其双胚苗性状有其胚胎学根源,双胚可能有4种来源,即来源于由双套胚囊形成的双胚、由多卵卵器中的卵细胞和类卵细胞分别通过受精作用形成双胚、由特异性的反足细胞通过异常发育后形成额外的幼胚、由胚乳细胞通过异常发育后形成胚乳型幼胚。通过对试验材料的受精频率与其结实率进行比较后发现,双胚苗水稻具有更弱的有性生殖能力,其遗传可塑性更强。
关键词 同源四倍体双胚苗水稻;激光共聚焦显微技术;雌雄配子体;发育特征
大量的研究结果表明,在植物界存在着两种具有潜在利用价值的优势效应,即杂种第一代所表现的杂种优势效应和通过染色体组多倍化所导致的优势效应[1]。根据杂交水稻育种的战略设想[2,3],在我国超级稻育种已经取得突破性研究进展并且在生产上也产生了巨大的经济效益的前提下,研究者有必要注重水稻遗传改良中仍然遗留下来的难点问题,即固定水稻杂种优势效应和简化水稻杂种优势利用的技术程序。如何利用现代生物技术创造水稻新种质,进而从生殖生物学角度寻找具有特殊生殖发育特性的新材料或新的基因资源则是值得进一步探索的研究方向。从禾本科植物的系统发育和演化历程来看,染色体组多倍化有助于改变原有物种的特征特性、增强新物种的适应性和提高其产量潜力。当二倍体水稻被诱导为同源四倍体水稻之后,后者的特征特性与前者的特征特性完全不同,这有可能为水稻遗传改良寻找到一条值得进一步探索的新途径。我们利用低能离子束生物技术对同源四倍体水稻进行注入处理后在其后代群体内筛选到一株具有双胚苗形态特征的同源四倍体突变株,经过多代的单株筛选和纯化之后已经证实,其双胚苗特性在群体内按照一定的频率表达,具有一定的遗传稳定性[4]。本项研究利用激光共聚焦显微技术对该双胚苗水稻的雄配子体和雌配子体的发育特征作了观察鉴定,旨在阐明其生殖发育特征,为进一步挖掘其潜在价值提供参考资料。
1 材料与方法
在对雄配子体的发育特征进行观察鉴定的试验中所采用的试验材料包括同源四倍体多胚苗水稻品系ASDOR05-01(图1)、同源四倍体水稻品系99-01(4)和二倍体水稻品系99-01(2)。以二倍体水稻品系99-01(2)为基础,通过种芽诱导法获得了同源四倍体水稻品系99-01(4)。随后,以同源四倍体水稻品系99-01(4)为材料利用低能氮离子束注入后经过多代筛选,获得了同源四倍体多胚苗水稻品系ASDOR05-01[4,5]。

图1 具有双胚苗特征的同源四倍体水稻
在对雌配子体的发育特征进行观察鉴定的试验中所采用的试验材料包括具有双胚苗特征的同源四倍体水稻株系ASDOR05-01。同时,以相应的两份亲本材料[即同源四倍体水稻品系99-01(4)和相应的二倍体水稻品系99-01(2)]作为对照。
在试验中利用I2-KI染色法观察试验材料成熟花粉粒的形态特征,由此确定其育性状况。在每份试验材料主穗上按照上部、中部和下部的不同位置分别取颖花作为试验材料。将颖花内的花药取出后放在载玻片上,利用浓度为0.1%的I2-KI溶液进行3~5min染色,在放大100倍的条件下对其进行镜检。根据败育花粉的表现形态以及对I2-KI溶液的染色反应将其分为典败、圆败、染败和正常等4种花粉粒。同时,对试验材料花粉粒的大小进行测定。以成熟花粉粒为材料,通过整体染色透明后将其放大200倍,在激光扫描共聚焦显微镜下测量花粉粒直径的大小,每份材料测定50粒花粉粒的直径,最后求取平均值。随后,通过稻穗进行套袋隔离后检查了试验材料的自交结实率。
在试验中采用整体染色透明法观察试验材料雄配子体的发育过程及其特征。在各份试验材料进入生殖生长之后,根据小孢子发育的不同时期与幼穗发育的形态特征进行取材,将小穗固定于FAA固定液(50%酒精∶冰醋酸∶甲醛=89∶5∶6)中,每份材料的每个发育时期取100个小穗。观察前用70%的酒精冲洗,然后在解剖镜下挑出花药,将其保存于70%酒精中备用。
在进行染色之前,首先将花药捣碎,由此将小孢子挤出,利用浓度分别为50%、30%、15%的乙醇对其进行梯度处理,每一梯度处理的时间为20min,再用蒸馏水冲洗。接着,利用浓度为1%的曙红Y溶液对试验材料进行8h的衬染,加入蒸馏水冲洗,换洗数次直至材料无浮色为止。随后,利用柠檬酸-磷酸氢二钠缓冲液(0.1mol/L,pH = 5.0)对试验材料进行预处理(8h左右),再利用浓度为20 μg/mL的H33342(Hoechest stain)溶液在25℃和黑暗条件下对其进行24h染色。利用蒸馏水冲洗2~3次,再利用不同浓度的乙醇(15%、30%、50%、70%、85%、95%)对其进行梯度脱水,每一次梯度脱水的时间为20min。再将染过色的试验材料放入无水乙醇中进行两次脱水,每次脱水的时间为2h,再转入无水乙醇中过夜留用。第2天将已经脱水的试验材料放入由无水乙醇和水杨酸甲酯(1∶1)组成的混合液中过渡1h,再利用水杨酸甲酯进行3次透明处理,在前2次透明处理中每次处理的时间为2h,最后1次透明处理的时间为15h。试验材料经过水杨酸甲酯透明处理后可以在水杨酸甲酯中保存备用。在进行观察之前,吸取一定量的处理好的试验材料将其置于载玻片上,加上盖玻片。将制备好的载玻片倒置于Leica SP2激光共聚焦扫描显微镜上,利用波长为405nm或488nm的激光对试验材料进行激发,通过电脑控制系统对试验材料进行扫描处理,由此获得小孢子在各个发育时期的外部形态和内部结构的连续性扫描图像。通过对试验材料的扫描图像资料进行分析后确定其小孢子发育的特异性。
在试验中采用细胞核荧光染色技术和子房整体透明技术对试验材料进行前处理。根据水稻的生育特性,在试验材料处于胚囊发育的8个时期分别取位于幼穗中部的颖花为试验材料,将其固定于FAA固定液中。在胚囊发育的每个时期获取100个颖花作为试验材料。在开始试验之前,首先利用70%的酒精将颖花冲洗一下,然后在解剖镜下从中分离出子房,将其保存在70%酒精中备用。在对试验材料进行染色之前,各个时期的子房材料分别经过浓度为50%、30%、15%的乙醇浸泡后将其放入蒸馏水中,乙醇浸泡的每一次梯度时间为20min。接着,在温度为60℃的水浴锅内利用1mol/L的HCl对试验材料水解15min。再利用1%的曙红Y对试验材料衬染8h。加入蒸馏水进行冲洗,换洗数次至无浮色为止。过后,利用柠檬酸-磷酸氢二钠缓冲液(0.1mol/L,pH = 5.0)对试验材料预处理8h,在温度为25℃的黑暗条件下利用20 μg/mLh33342(Hoechest stain)对其进行染色24h。利用蒸馏水对试验材料冲洗2~3次,利用乙醇梯度(15%、30%、50%、70%、85%、95%)进行脱水,每一级的脱水时间为20min。随后,将子房放入无水乙醇中脱水两次,每次脱水的时间为2h,再将其转入无水乙醇中过夜。第2天将已经脱水的材料放入由无水乙醇和水杨酸甲酯(1∶1)组成的混合液中过渡1h,再用水杨酸甲酯透明处理3次。在前两次透明处理中每次处理的时间为2h,最后一次透明处理的时间为15h。试验材料经过水杨酸甲酯透明处理后可以在水杨酸甲酯中保存备用。在对试验材料进行观察之前,用镊子轻轻将处理好的子房夹出,将其置于凹玻片上,利用丁香油进行封片处理。将制备好的载玻片倒置于Leica SP2激光扫描共聚焦显微镜上,利用波长为488nm的激发光对试验材料进行激发处理,通过激光扫描后可以获得胚囊各个发育时期的清晰图片。通过对图像资料进行分析、比较和综合之后确定试验材料在胚囊发育的各个时期所表现的特异性。
在试验中对3份试验材料的雌配子体的育性与其结实率的相关性进行了比较研究。在田间条件下按照顺序排列方式进行单株种植,每份试验材料的群体为200株,采用常规的田间管理措施。在试验材料始穗后30 d从每份试验材料的群体内随机地取10株材料进行考种。通过对考种结果进行比较分析后确定其结实率特点以及试验材料的育性与其结实率的相关性。
2 结果与分析
2.1 雄配子体的发育特征
关于同源四倍体水稻雄配子和雄配子体的发育特征,前人已经有过一些研究报道[6,7]。经过染色体组多倍化之后将导致多倍体水稻后代表现出更弱的有性生殖能力,在遗传上其可塑性增强而保守性减弱;在小孢子发育上正常雄配子或雄配子体的数量减少而败育雄配子或雄配子体的数量增加。本试验的分析结果表明,3份试验材料在成熟花粉粒的育性和结实率上存在着明显的差异,在其花药内均包含有正常花粉粒和败育花粉粒,败育花粉粒包括典败型、圆败型和染败型。在同源四倍体双胚苗水稻品系ASDOR05-01的花药内正常花粉粒数量最少而败育花粉粒数量最多,其结实率最低。在同源四倍体水稻品系99-01(4)的花药内情况居中。在二倍体水稻品系99-01(2)的花药内正常花粉粒数量最多而败育花粉粒数量最少,其结实率最高(表1)。由此可见,在每份试验材料中均存在着正常雄配子体和异常雄配子体,而同源四倍体双胚苗水稻品系ASDOR05-01表现出更弱的有性生殖能力。除此之外,通过比较各份试验材料的花粉粒形态后发现,在成熟花粉粒的大小上同源四倍体水稻均比二倍体水稻要大一些,在两份同源四倍体水稻之间其成熟花粉粒的大小很接近,不存在着明显的差异(表2),这些观察结果与前人所报道的结果相吻合[1]。
表1 成熟花粉粒的育性(%)与结实率(%)
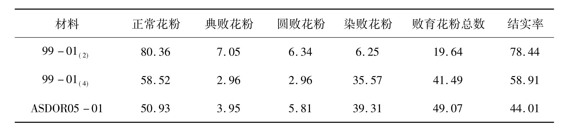
表2 成熟花粉粒的大小

在试验中对正常雄配子体的发育特征进行观察鉴定的结果值得注意。在3份试验材料中,正常雄配子体的发育过程和特征基本相似,没有表现出因试验材料的种类所呈现出的明显差异。在小孢子母细胞形成期,小孢子母细胞为近圆形,其体积比花药内其余细胞的体积都要大一些,尤其细胞核比较大,细胞质比较浓厚。小孢子母细胞的细胞壁为薄薄的一层,染色很浅。在小孢子母细胞减数分裂期,小孢子母细胞的形状由近圆形慢慢地变成椭圆形。当小孢子母细胞中营养物质积累达到一定程度时,就开始进行减数分裂。在前期Ⅰ的情况比较复杂。在细线期,小孢子母细胞的细胞核和核仁体积明显增大,染色比较深,在透明的细胞核液中出现细长的染色质丝。染色体开始显现为丝状,互相交错地布满于整个细胞核内,细胞质比较浓。在偶线期,小孢子母细胞的两条同源染色体进行配对,染色体呈极化状态,即在染色体靠近细胞核膜的一端集中成束,而在其另一端散成花朵状。在粗线期,小孢子母细胞内的染色体逐渐缩短变粗,每条同源染色体发生纵裂为2条染色单体,由着丝点将其连接在一起,此时每一对同源染色体具有4条染色单体。在双线期,小孢子母细胞的染色体进一步缩短变粗,发生配对的同源染色体在一部分区域分开,在另外的部分区域发生联合交错互换,整个双价染色体呈现出多种交错形状。在终变期,小孢子母细胞内的染色体极端缩短成颗粒状,同源染色体成对地排列在细胞核的内膜周围。在终变期末期,纺锤丝出现,核仁和核膜消失,减数分裂前期Ⅰ结束。在中期Ⅰ,双价染色体排列在赤道板上,每对同源染色体中两条染色体的着丝点向着相对的两极。在激光扫描共聚焦显微镜下能清晰地看到染色比较浅的纺锤体和染色比较深的染色体整齐地排列在赤道板位置上。在后期Ⅰ,每一对同源染色体在着丝点位置发生分离后向两极移动,但每条染色体仍然包含着已经发生部分区段的纵列而未完全分开的2条染色单体。在末期Ⅰ,染色体达到纺锤体的两极后浓缩成团。在此时,核膜和核仁重新出现并形成2个子细胞核,在每一个子细胞核内染色体数目比花粉母细胞内的染色体数目减少一半。随后,在赤道板上产生成膜体并形成新的细胞壁,由此将其分成2个子细胞,即形成二分体。在减数分裂Ⅱ期间,染色体的动态变化与一般有丝分裂期间染色体的动态变化很相似,最终由2个子细胞形成4个子细胞,四分体的出现标志着减数分裂过程的结束。在小孢子发育时期及其以后的各个时期,3份试验材料均表现出一些共性。在四分体中的每一个小孢子进一步发育成外观形状为圆形的单核细胞,其细胞壁周围的胼胝质被溶解之后,每个细胞彼此分开,即进入小孢子早期。小孢子在早期表现为圆球形,细胞体积比较小,其细胞壁仍然很薄,在细胞质中无液泡,细胞核比较大,位于细胞的中央。在小孢子中期,其体积明显增大,细胞壁显著加厚并出现萌发孔,在细胞质中出现液泡但尚未形成中央大液泡。在小孢子晚期,中央大液泡形成,细胞核的体积增大,小孢子的壁增厚。在此期间,小孢子的细胞核均被大液泡挤至细胞的边缘,即单核靠边期。在二胞花粉期,小孢子经过第一次有丝分裂后形成2个细胞,即形态上比较大的营养细胞和比较小的生殖细胞。生殖细胞没有细胞壁结构,紧挨着营养细胞的细胞壁存在。在二胞花粉早期,在小孢子内还没有淀粉粒的积累而在二胞花粉晚期,在小孢子内开始出现淀粉粒。淀粉粒在细胞质中不断积累,液泡的体积逐渐缩小,并被推至萌发孔的对侧。生殖细胞与花粉壁脱离,逐渐移至营养核附近。在成熟花粉期,在花粉粒内充满了淀粉粒等贮藏物质,花粉粒形态为圆球形,生殖细胞完成第二次分裂,由此形成两个精细胞。由于在成熟花粉粒内充满了淀粉粒等物质,利用一般的染色方法很难观察到其内的3个细胞核。本项研究中,利用特异性的核荧光染料进行染色,在激光扫描共聚焦显微镜下可以清晰地观察到3个细胞核的存在,2个生殖细胞核的形态呈梭形,营养细胞核呈圆形。
在试验中对异常雄配子体的败育特征进行观察鉴定的结果也值得注意。通过对3份试验材料进行比较后发现,同源四倍体双胚苗水稻品系ASDOR05-01表现出更加明显的特异性。在小孢子母细胞进行减数分裂时,出现了5种不正常的分裂现象,即部分染色体不能正常配对、在分裂中期形成了三极纺锤体、形成异常的二分体、在第二次减数分裂中期出现了分裂不同步现象(即在二分体中1个子细胞处于分裂中期而另一个子细胞此时已进入分裂后期)、形成异常的四分体(即形成的四个子细胞不能正常分开,在细胞之间存在着不完全的细胞板将其连在一起)。在单核小孢子期同源四倍体双胚苗水稻品系ASDOR05-01的部分小孢子便发生败育,由此形成典败花粉粒。观察结果表明,部分小孢子发生败育的方式有多种,有的败育花粉粒由于小孢子内的细胞质发生浓缩,花粉壁出现皱折变形所致;有的败育花粉粒因小孢子内的细胞质破碎为块状而导致其发生败育;有的败育花粉粒起因于小孢子的内含物完全消失后形成空泡状结构;还有的败育花粉粒来源于小孢子细胞发生变形,由此形成弯月形结构,其内的细胞核仍然清晰可见。在二胞花粉早期同源四倍体双胚苗水稻品系ASDOR05-01的部分小孢子发生败育后,形成圆败花粉粒。观察结果表明,二胞花粉的败育始于生殖核发生解体。在这种情况下,在小孢子中出现比较多的染色质团块,生殖核发生解体之后促使营养核也开始解体,由此也伴随着染色质团块的出现。由生殖核和营养核所产生的染色质团块聚集在一起之后,在二胞花粉中形成巨大的染色质团块。经过核物质染料H33342染色后在二胞花粉内可以见到染色比较深的染色质团块。随后,在二胞花粉内的这些染色质团块随着发育进度的推进而逐渐消失,最后在花粉粒内很难寻找到内含物,仅剩下空壳花粉粒。在二胞花粉晚期或三胞花粉早期同源四倍体双胚苗水稻品系ASDOR05-01的部分小孢子发生败育后,形成染败花粉粒。观察结果表明,染败发生在二胞花粉晚期或三胞花粉早期。在此期间,在花粉粒内存在着淀粉粒积累现象但淀粉粒尚未完全充满,生殖细胞还没有发生分裂,反而逐渐解体和消失。在二胞花粉晚期,可以明显地观察到2个细胞核的存在,并且在花粉粒中已经有淀粉粒的积累。有的染败花粉粒处于三胞期,在显微镜下3个细胞核明显可辨,只是其中的内含物比较少。(www.chuimin.cn)
2.2 雌配子体的发育特征
在本试验的观察鉴定中已经确定,对于正常雌配子体的发育特征而言,所有的试验材料均表现出明显的相似性。在孢原母细胞形成期,在珠心组织的表皮下会分化出形态上具有一定特征的孢原细胞,即孢原细胞在形态上呈近圆形,体积比较大,细胞质比较浓厚,细胞核比较明显,在细胞核内具有一个染色比较深的核仁。在大孢子母细胞形成期,随着内珠被细胞和外珠被细胞不断伸长的同时,孢原细胞也明显地伸长和增大,由此分化出大孢子母细胞。大孢子母细胞的细胞核和核仁比较明显,在其细胞核内存在着染色比较深的染色质。在此时,胚珠逐渐发生倒转生长。在大孢子母细胞减数分裂期,大孢子母经过第一次分裂后形成二分体。二分体经过第二次分裂后形成四分体。在功能大孢子形成期,在四分体中靠近珠孔端的3个大孢子依次发生退化,即珠孔端的大孢子最先退化,然后是靠珠孔端的第2个和第3个大孢子相继发生退化,最后只有近合点端的大孢子经过发育后形成功能性大孢子。在单核胚囊形成期,功能大孢子的体积逐渐增大,在细胞内开始出现液泡,其细胞核偏向于珠孔端。在胚囊有丝分裂期,在单核胚囊内的细胞核经过1次有丝分裂后形成两个无壁的细胞核。在细胞骨架系统的牵引下,无壁的细胞核分别移向胚囊的珠孔端和合点端,由此形成2核型胚囊。在此时,在胚囊中间存在着一个比较大的液泡。在2核胚囊内,位于珠孔端和合点端的细胞核分别发生2次连续分裂,由此分别形成4核胚囊和8核胚囊。在正常情况下,胚囊内的有丝分裂会同步进行,即由单核胚囊发生分裂后会分别形成2核胚囊、4核胚囊和8核胚囊。在8核胚囊发育期,首先由珠孔端和合点端的1个细胞核发生位移,即分别移向胚囊的中部,由此形成中央细胞的2个极核。在此时,胚囊内的各个细胞核逐渐地发生细胞化,即在合点端的3个细胞核分别发育为3个反足细胞;在珠孔端的3个细胞核分别发育为1个卵细胞和2个助细胞。在成熟胚囊期,在卵器中的1个助细胞已经发生退化,在成熟胚囊内往往只能见到1个助细胞。助细胞的细胞核比较小,具有1个比较小的核仁。在助细胞的珠孔端存在着具有特异性的丝状器。卵细胞呈梨形,位于珠孔端部分比较小而位于合点端部分比较大。其细胞核往往位于近珠孔端,其中有1个比较大的、染色比较深的核仁。在卵细胞的合点端内存在着1个大的液泡。在成熟胚囊中,卵细胞的细胞质一般比较稀薄,在发生受精作用之前,仅仅在其细胞核的外围形成比较浓厚的细胞质。中央细胞为1个高度液泡化的细胞,其形状比较大。2个极核紧靠在一起,在每1个极核内各存在着1个比较大的核仁。2个极核在发生受精作用之前并不会融合成次生极核。反足细胞位于胚囊内的合点端,在发生受精作用之前反足细胞通过分裂后已经增殖为多个细胞,在每个细胞内往往存在着多个形态上不规则的细胞核。反足细胞的染色质浓厚,染色往往比较深。
在试验材料雌配子体的发育过程中已经观察到一些异常现象。观察鉴定结果表明,在雌配子体发育过程中同源四倍体水稻品系比相应的二倍体水稻品系出现异常胚囊的比例更大。在两份同源四倍体水稻品系中,双胚苗品系ASDOR05-01比其亲本99-01(4)表现出更加明显的特异性。在ASDOR05-01植株的雌配子体发育过程中,其特异性主要包括11种类型。其一是在减数分裂Ⅱ期间发生子细胞分裂不同步现象。在二分体时期,当位于近珠孔端的一个子细胞已经进入分裂后期,而近合点端的一个子细胞仍然没有启动新的细胞分裂。其二是在四分体时期,四个子细胞全部发生退化,以至于胚珠内只剩下一个空腔而不存在胚囊结构。其三是在胚囊内发生有丝分裂的过程中,细胞核的数目及其分布出现异常。在观察中发现,在1个胚囊内存在着6个细胞核,其中珠孔端4个,合点端2个,并且在胚囊内所有细胞核几乎呈直线分布。其四是胚囊的构型异常。在有的胚珠内珠孔端外部有一列呈直线排列的细胞,由此将胚囊挤在很小的空间里。在胚囊的珠孔端无卵器结构,胚囊内只存在着3个反足细胞和2个极核。其五是在胚囊内极核数目或位置发生异常。在胚囊发育过程中,两个极核均发生退化,以至于在胚囊内没有极核存在;或者在中央细胞内1个极核发生退化,以至于在成熟胚囊内只存在着1个极核;在有些成熟胚囊内极核没有正常地移位至卵器的上端,而是位于胚囊的中央,或位于合点端,或靠近反足细胞团。其六是在胚囊内的卵器发生退化或卵器细胞出现异常。在胚囊内,有的卵器发生退化后在卵器的位置上不存在正常的细胞结构;在有的成熟胚囊中卵器内的两个助细胞全部退化,以至于在卵器内只存在着1个卵细胞。其七是雌性生殖单位发生退化。在有些胚囊内,卵器和极核均发生退化,以至于在胚囊内只能观察到反足细胞团。其八是胚囊在发育过程中逐渐发生解体,最终导致胚囊退化。其九是子房和胚囊细胞在发育过程中发生退化,试验材料经过染色后细胞壁显示出强烈的荧光。其十是在同一个胚珠内出现多卵卵器胚囊,这很可能是形成多胚的原因之一。其十一是出现双套胚囊现象。在观察中发现,在同一个胚珠内出现了双套胚囊现象,两个胚囊相对分布并且各有一套卵器,一个胚囊的卵器位于珠孔端,而另一个胚囊的卵器位于合点端(图2)。在试验中通过对ASDOR05-01的98个成熟胚囊进行观察鉴定后确定,其中64个胚囊具有正常的胚囊结构,其正常胚囊的频率为65.31%;在34个胚囊内出现了退化现象或异常现象,其异常胚囊的频率为34.69%。

图2 在ASDOR05-01材料中所存在的双套胚囊
在试验中对试验材料成熟胚囊的结构与其育性的关系进行了比较研究。研究结果表明,在所有试验材料中成熟胚囊的结构状态与其育性表现存在着一定的相关性。在二倍体水稻中正常胚囊所占的比率比较高,而异常胚囊的频率比较低,其结实率也比较高。同源四倍体水稻与二倍体水稻相比,前者的异常胚囊的频率明显增加,其结实率也显著下降。在两份同源四倍体水稻材料中,ASDOR05-01的异常胚囊的频率比较高,其结实率比较低(表3)。由此可见,经过离子束注入处理后所筛选到的双胚苗水稻材料在生殖发育特性上表现出更加明显的特异性,即表现出更弱的有性生殖能力。
表3 成熟胚囊的类型与其结实率

3 讨论
从水稻遗传改良的现状来看,其研究难题之一就是在确保水稻产量潜力的前提下如何简化利用水稻杂种优势的技术程序,即固定水稻的杂种优势。我国水稻无融合生殖的研究主要集中在筛选和鉴定多胚苗水稻,对于多胚苗水稻的形态学、胚胎学和遗传学的研究已经获得了一些具有学术价值的研究资料[8]。然而,前人关于多胚苗水稻的研究主要在二倍体水平展开探索,而关于同源四倍体多胚苗水稻的研究目前尚未见到公开的文献报道。根据禾本科植物中无融合生殖物种的特异性,在多倍体水平寻找水稻无融合生殖种质则成功的可能性比较大[1,9]。
本项研究的结果表明,在每份试验材料中均存在着正常雄配子体和异常雄配子体,而同源四倍体双胚苗水稻品系ASDOR05-01表现出更弱的有性生殖能力。在3份试验材料中,正常雄配子体的发育过程和特征基本相似,没有表现出因试验材料的种类所呈现出的明显差异。同源四倍体双胚苗水稻品系ASDOR05-01在小孢子母细胞进行减数分裂时,出现了5种不正常的分裂现象,即部分染色体不能正常配对、在分裂中期形成了三极纺锤体、形成异常的二分体、在第二次减数分裂中期出现了分裂不同步现象(即在二分体中1个子细胞处于分裂中期而另一个子细胞此时已经进入分裂后期)、形成异常的四分体(即形成的四个子细胞不能正常分开,在细胞之间存在着不完全的细胞板将其联结在一起)。在花粉发育过程中雄配子或雄配子体所表现出的比较高的败育频率可能是导致其结实率比较低的主要原因之一。同源四倍体双胚苗水稻品系的雄配子体发生败育的生物学现象是一个相当复杂的代谢过程,小孢子在各个发育时期产生败育的细胞学机制还有待于进一步研究。
在对雌配子体的发育特征进行观察的过程中已经发现一些异常现象。观察鉴定结果表明,在雌配子体发育过程中同源四倍体水稻品系比相应的二倍体水稻品系出现异常胚囊的比例更大。在两份同源四倍体水稻品系中,双胚苗品系ASDOR05-01比其亲本99-01(4)表现出更加明显的特异性。在ASDOR05-01植株的雌配子体发育过程中,其特异性主要包括11种类型。同源四倍体水稻和二倍体水稻在发生受精作用的时间上没有存在着很大的差异。双胚苗水稻的受精作用和胚胎发育表现出明显的特异性,其双胚苗性状有其胚胎学根源,双胚可能有4种来源,即来源于由双套胚囊形成的双胚、由多卵卵器中的卵细胞和类卵细胞分别通过受精作用形成双胚、由特异性的反足细胞通过异常发育后形成额外的幼胚、由胚乳细胞通过异常发育后形成胚乳型幼胚。通过对试验材料的受精频率及其结实率进行比较后发现,双胚苗水稻具有更弱的有性生殖能力,其遗传可塑性更强,这为离子束介导技术的研究找到了比较好的受体材料。除此之外,在所有试验材料中成熟胚囊的结构状态与其育性表现存在着一定的相关性。
[1]黄群策.被子植物的无融合生殖[M].福州:福建科学技术出版社,2000.
[2]袁隆平.杂交水稻的育种战略设想[J].杂交水稻,1987,2(1):1-3.
[3]袁隆平.杂交水稻超高产育种[J].杂交水稻,2001,16(1):1-3.
[4]黄群策,梁秋霞,李玉峰,等.低能离子注入同源四倍体水稻的生物学效应[J].激光生物学报,2003,12(5):355-359.
[5]黄群策,代西梅.低能氮离子对不同倍性水稻的诱变效应[J].杂交水稻,2004,19(3):57-61.
[6]黄群策,孙敬三,白素兰.同源四倍体水稻的生殖特性研究[J].中国农业科学,1999,32(2):14-17.
[7]黄群策,代西梅.水稻同源多倍体的研究策略[J].杂交水稻,2006,21(2):1-4.
[8]郭学兴.中国水稻无融合生殖研究进展[M].成都:四川科学技术出版社,1991.
[9]黄群策,秦广雍.禾本科植物染色体组多倍化研究[M].北京:原子能出版社,2008.
有关水稻染色体组多倍化研究的文章

同源四倍体双胚苗水稻雌配子体的发育特征黄群策代西梅贾宏汝摘要利用激光扫描共聚焦显微技术对3份水稻材料的雌配子或雌配子体的发育特征进行了观察鉴定。2结果与分析2.1试验材料正常雌配子体的发育特征根据前人的研究结果,水稻雌配子体的正常发育过程有其特点。......
2023-11-28

第二篇关于同源四倍体水稻的诱导文献综述根据进化论的观点,植物物种的形成和分化要经过漫长的历程,而不同物种在演化过程中所经历的艰难程度都大不相同。其二,在种子露白时进行诱导处理只能使胚芽鞘明显膨大而不能从其后代中筛选到四倍体材料;先行预处理,使胚芽鞘伸长到2.0cm左右时再用秋水仙素对种芽进行诱导处理则能从其后代中筛选到四倍体材料。......
2023-11-28

同源四倍体水稻的生殖特性研究黄群策孙敬三白素兰提要与相应的二倍体水稻相比,同源四倍体的生殖特性发生了明显的变化,有性生殖能力明显变弱。同源四倍体水稻有性生殖的异常可能为无融合生殖基因的表达创造了条件。由此可见,同源四倍体水稻的雄配子体的育性已经明显降低,这是其有性生殖能力明显变弱的表现之一。......
2023-11-28
从低能离子束生物工程的发展趋势来看,对于突变体材料所表现出的后效性值得关注,因而也对于同源四倍体双胚苗水稻的生殖发育特性值得进行深入研究。关于二倍体水稻的双胚苗的形态特征及其生殖发育特性,前人已经进行过一些研究[4-6],而关于同源四倍体双胚苗水稻的雄配子体发育特性尚未见到公开的文献报道。具体见上篇“同源四倍体双胚苗水稻的生殖发育特征研究”。......
2023-11-28
在同源四倍体水稻中的子房膨大效应比二倍体水稻中的子房膨大效应更加明显。本文报道了以多个同源四倍体水稻为杂交母本与狼尾草进行杂交,由此获得实粒种子的试验结果。相反,同源四倍体水稻的有性生殖能力比较弱,其遗传保守性比较小而遗传可塑性比较大,利用同源四倍体水稻与狼尾草进行杂交则难度比较小,成功的可能性比较大。......
2023-11-28

由此认为,同源四倍体水稻的有性生殖能力已经明显变弱。关于二倍体水稻的受精过程的细胞学观察,前人已有报道[5-7]。但是,关于同源四倍体水稻的双受精过程,目前国内外尚未见到公开的文献报道。对二倍体水稻和同源四倍体水稻的双受精过程进行比较研究将有助于了解其生殖特性的差异,这对于从同源四倍体水稻中鉴定和筛选潜在的无融合生殖基因具有重大的现实意义。......
2023-11-28
双胚苗水稻种子的扫描电镜观察黄雅琴黄群策燕晓阳摘要以同源四倍体双胚苗水稻品系及其衍生后代为研究材料,利用扫描电镜技术和X射线衍射技术对稻米的垩白性状淀粉粒的形态特征和淀粉的晶体特性进行了初步研究。关键词双胚苗水稻;同源四倍体;米质;淀粉粒;扫描电镜; X射线衍射仪水稻是世界上最主要的粮食作物,选育高产、优质、多抗统一的水稻新品种是21世纪的育种目标[1]。......
2023-11-28
其一,氮离子注入后在同源四倍体水稻的后代群体内可以筛选到具有不同特征特性的突变单株,利用离子束生物技术对同源四倍体水稻进行遗传改良的技术路线值得肯定,其实用性已经为同源四倍体水稻新种质的选育找到了新的突破口。......
2023-11-28
相关推荐