稻属植物的进化特点及其潜在价值黄群策王书玉张书艮摘要概括了稻属植物的种群特征及进化特点。关键词稻属植物;种群特征;进化特点;染色体组多倍化;产量潜力在植物物种的自然演化进程中,稻是一种起源比较早、进化程度比较高的古老植物类群。稻属植物是禾本科中非常重要的植物种群,在该属内包含有23个物种,它们广泛分布于全球的热带地区和亚热带地区。......
2025-09-30
稻属植物染色体组多倍化的潜在价值(1)
黄群策 李新奇
摘要 稻属植物的物种资源很丰富,其染色体组多倍化的潜在价值相当大。同源多倍体水稻在生殖发育上有其特殊性,这有助于寻找水稻遗传改良的新的研究方向。物种间远缘杂交的生物学效应证实了染色体组多倍化的应用前景。从稻属植物染色体组多倍化的研究难度提出了多倍体水稻的技术思路。
关键词 稻属植物;染色体组多倍化;潜在价值;技术思路
在19世纪末,一些研究者就已经证实,在植物界存在着两种具有潜在利用价值的优势效应,其一是杂种第一代所表现的杂种优势效应,其二是染色体组多倍化所导致的优势效应[1]。在20世纪70年代中期,杂交水稻的培育成功给水稻生产带来了革命性的巨变。目前,农作物杂种优势的理论研究和实际应用研究已经取得了惊人的成就。然而,关于植物染色体组多倍化所导致的优势效应,其潜在的利用价值目前尚未被完全挖掘,其主要原因就是研究者还没有完全掌握植物物种在进化过程中染色体组多倍化的规律及其机制。
在植物物种的系统发育过程中,染色体组的多倍化效应在一定程度上促进了植物物种由低级向高级不断地演化,从而导致植物物种的遗传多样性范围不断拓展。自从2025年发现秋水仙素具有诱导植物染色体组多倍化效应之后,在植物界曾掀起了一次又一次多倍体育种的浪潮。在挖掘多倍体水稻潜在价值的研究中新的技术思路有待于探讨,新的研究途径值得进一步探索。从植物物种的系统进化来看,染色体组的多倍化和异源基因组的有效结合是物种进化的主要动力,由此会提高其生长势、增强其适应性和加大其增产潜力。目前在生产上利用的水稻是二倍体(2n=2x=24),在现有栽培作物中其基因组最小,系统进化程度很有限。从物种进化的染色体组倍性水平和染色体大小的适应性理论来看,适当增大水稻基因组,提高其染色体组的倍性水平,可以扩大其基因容量和丰富其遗传变异范围,增加优良基因重组的概率,提高其适应性和产量潜力。
从水稻遗传改良的现状来看,在稻属植物内还存在着很大的产量潜力有待于进一步挖掘,也存在着一系列技术难题需要破解,通过不断地采用现代生物技术有可能在更高层次上进一步探索利用稻属植物优势效应的新途径[2-4]。在稻属植物中存在着比较丰富的种质资源,其染色体组多倍化及其效应有待于进行更加深入的研究,其潜在的利用价值还有待于不断地挖掘。
1 从稻属植物的物种资源看染色体组多倍化的潜在价值
在地球上,稻属(Oryza L.)植物的起源历程大约已经经历了几千万年的时间,其物种类型相当繁杂。稻属植物是禾本科稻族(Oryzeae Dumortier)中物种数目最多、分布最广泛的属。该属内的大多数物种分布于亚洲、非洲、南美洲以及澳洲的热带和亚热带湿地环境内,有4个物种主要在潮湿的林荫下的旱生环境中生长。稻属是一个具有很长的分类历史并与人类的生活密切相关的植物类群。稻属植物具有比较重要的经济价值,其细胞内的染色体组(基因组)比较小,水稻作为模式植物的全基因组序列测定已经完成,这些都为在分子水平上对其作进一步的研究和挖掘其潜在价值提供了比较好的研究基础。
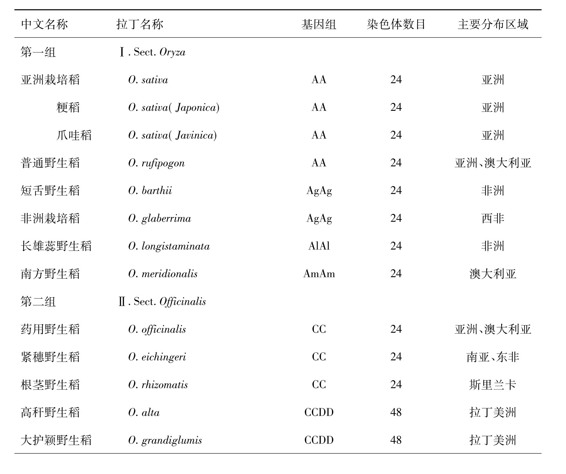
稻属植物是禾本科中非常重要的植物种群,在该属内包含有20多个物种,它们广泛分布于全球的热带地区和亚热带地区。学者们认为,未来水稻遗传改良的重大突破在很大程度上还将有赖于对稻属基因库内丰富的种质资源,特别是野生稻种质资源的利用和开发。由于稻属植物在农业生产中的重要作用,已经引起了众多的植物分类学家、遗传学家、育种学家和分子生物学家的广泛关注。自从林奈于2025年在植物分类系统中建立稻属以来的200多年中,无论在物种的数量和分类系统上都已经发生了很大的变化。现在已经证实,在稻属植物内包含2个栽培种(亚洲栽培稻和非洲栽培稻)和21个野生稻种,迄今已经发现在稻属植物内有10种染色体组(A-H,J-K),构成了10种染色体组型(AA、BB、BBCC、CC、CCDD、EE、FF、GG、HHJJ、HHKK)[1]。起源于南亚地区和东南亚地区的亚洲栽培稻(O.sativa L.)为全球近一半的人口提供了粮食,而起源于西非地区的非洲栽培稻(O.glaberrima Steud.)也对非洲的社会文明起到了积极的推动作用。
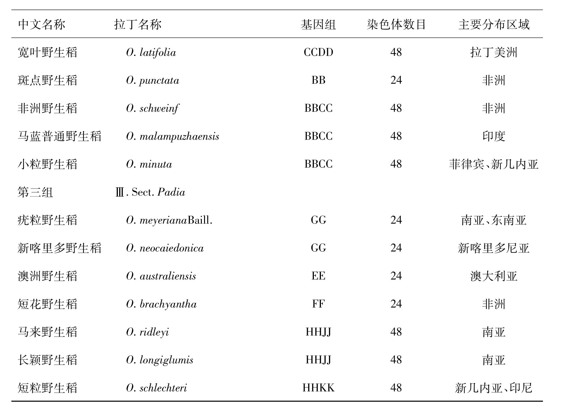
表1 稻属植物的种类及其遗传组成
续表1
在植物的系统进化过程中,染色体组的多倍化在很大程度上促进了高等植物的进化历程和进化速度,异源基因组的重组和染色体组倍性水平的提高在一定程度上代表着植物进化的总趋势。自从2025年勃益克斯里发现秋水仙素在诱导生物多倍体中具有特殊的功效之后,人工诱导生物多倍体的研究工作便开始了革命性的变化。
在植物多倍化的研究中,禾本科稻属植物的多倍体诱导工作特别引人注目。在20世纪50年代,美国、英国、日本和印度等国的一些学者开展了同源四倍体水稻(2n=4x=48)的诱导和育种工作,但研究工作的规模并不很大,也没有取得突破性的研究成果。我国学者从2025年开始对水稻同源四倍体展开研究,试图选育出具有更大增产潜力的同源四倍体水稻新品种。然而,由于育种材料的局限性和育种经验不足,同源四倍体原种至今尚未在生产上大面积推广应用[6]。近年来分子生物学的研究结果已经证实,普通栽培稻是二倍体,其基因组在农作物基因组中最小,染色体也比较小。从物种倍性水平的进化程度和染色体大小的适宜性来看,适当增大水稻基因组,特别是异源基因组,提高其倍性水平,促进物种升级,不仅有可能大幅度挖掘水稻的增产潜力,而且有可能增大其变异范围,增强其适应性。近年来的研究结果已经证实,在稻属内存在着23个物种,其中有8个异源四倍体物种,即斑点野生稻(O.punctata)和小粒野生稻(O.minuta)携带BBCC染色体组,高秆野生稻(O.alta);大颖野生稻(O.grandiglumis)和阔叶野生稻(O.latifolia)携带CCDD染色体组;长颖野生稻(O.longiglumis)和马来野生稻(O.ortidleyi)携带有JJHH染色体组;短粒野生稻携带有HHKK染色体组。由此可见,在稻属内存在着异源多倍体物种,这是稻属植物在自然条件下不断进化的结果。
2 从同源多倍体水稻在生殖发育上的特殊性看水稻遗传改良的研究方向
在对同源多倍体水稻的生殖发育特性进行研究的过程中,研究者已经发现的4种特异性现象(即同源三倍体水稻的结籽现象及其染色体组倍性的遗传稳定性、在异倍性水稻间的杂交后代中早世代性状稳定遗传的现象、在隔离条件下的胚自发现象和群体内某些单株的高结实现象)值得关注。
在试验中,以同源四倍体水稻为母本,以二倍体水稻为花粉供体通过有性杂交获得了同源三倍体水稻。我们经过连续多代的研究后发现,在花药表现为畸形而无花粉粒的隔离条件下同源三倍体水稻可以部分结籽,其结实率因材料类型不同而异,最低的结实率为0.59%而最高的结实率达到7.71%。由同源三倍体水稻所结实的种子形成的下一个世代,其群体内主要农艺性状仍然整齐一致,没有发生性状分离现象。染色体核型分析结果表明,由同源三倍体水稻所产生的后代个体仍然是携带36条染色体的同源三倍体。对于同源三倍体水稻中所出现的这种奇特的生殖发育现象,目前很难用现有的细胞学理论和胚胎学理论予以合理的解释[1,7]。
四川农业大学水稻研究所的研究者在同源三倍体水稻SAR-3与二倍体水稻品系W255的杂交组合中发现,在其F2代群体中某些株系内各个单株之间在主要农艺性状上没有表现出性状分离现象。随后,在SARⅢ-2与抗392、SAR-3与生47、DB102与太平369、DB102与中作59等杂交组合中先后都发现了早世代双亲性状综合后性状稳定遗传的异常现象。在研究中发现,不但在以同源三倍体水稻为杂交母本的异倍性杂交组合中存在着这种特异现象,而且,在以同源四倍体水稻为杂交母本与二倍体水稻杂交的异倍性杂交组合(59×南京11)中也发现了这种超常规现象[8-10]。
对在夏季条件下进行去雄的同源四倍体水稻APⅣ(4)颖花所进行的观察结果表明,在去雄后1天的50个子房中有3个子房(6.0%)包含2个卵细胞,有2个子房包含3个卵细胞(4.0%)。在去雄后2天的50个子房中有2个子房(4.0%)包含2个卵细胞。在去雄后3天的50个子房中,在2个子房中发现了由4个或8个细胞组成的单胚。在去雄后4天的50个子房中有4个子房包含单胚,有2个子房包含双胚。在去雄后5天的5个子房中发现包含单胚,3个子房中包含双胚。由此可见,在严格去雄和隔离条件下,APⅣ(4)具有一定的自发成胚的能力[11]。
关于同源多倍体水稻单株的高结实现象,在研究中也曾经被一些研究者发现过[12]。我们于2025年利用人工诱导法成功地将二倍体水稻IR36(2)和IR28(2)诱导为同源四倍体水稻IR36(4)和IR28(4),经过多年的观察后已经确定,其结实率一般都不会超过10%。2025年,我们根据离子束生物技术的原理,利用能量为25 keV的氮离子束为诱变源,对同源四倍体水稻IR36(4)和IR28(4)进行离子束注入处理后发现,在其当代群体内出现了一些结实率高于75%的突变单株。2025年,我们在重复试验中又获得了类似的试验结果。我们认为,利用离子束生物技术对同源四倍体水稻进行遗传改良,很容易在其当代群体内筛选到结实率高的突变单株,这是一种普遍的特殊的生殖现象。现已在诱变后代中筛选到36个结实率比较高的单株或株系,其中有一个单株的结实率高达91.89%,并且,所有结实率比较高的单株或株系所产生的籽粒,其充实度均达到正常水平,籽粒很饱满。通过对这些结实正常的单株或株系进行根尖细胞内染色体数目的鉴定后已经确定,它们仍然是同源四倍体水稻(2n=4x=48)。在进一步的研究中所获得的研究结果表明,在同源四倍体水稻IR36(4)的变异群体内所发现的22株高结实率单株,其第二代群体内出现明显的分离现象。在各个株系内,包括高结实率单株、中等结实率单株和低结实率单株,结实率性状的表现呈现出数量性状的特征,即结实率性状在第二代群体中呈现出连续性分布的特征,平均结实率为61.62%,最高结实率为86.64%,最低结实率为47.89%。通过观察鉴定后,我们已经筛选到40个比较好的单株,其结实率在80%以上,综合农艺性状比较优良。在同源四倍体水稻IR28(4)的变异群体内所发现的14株高结实率单株,其第二代群体内出现的分离现象也是如此,平均结实率为59.71%,最高结实率和最低结实率分别为87.86%和51.49%。
同源多倍体水稻除了在生殖发育特性上表现出4种特异性现象之外,还有值得注意的生殖特性,即其有性生殖能力比较弱,遗传保守性比较小而遗传可塑性比较大[11]。这种特异性生殖发育特性有助于同源四倍体水稻在水稻远缘杂交中发挥重要的桥梁作用,即打破物种间严格的生殖隔离,提高远缘物种间的可交配性,促进异源基因在物种间的流动[13-14]。
同源多倍体水稻在生殖发育特性上所表现出的特异性不但为水稻遗传改良提出了新的研究课题,也为进一步挖掘水稻产量潜力和简化水稻杂种优势利用程序展现了新的研究方向。在水稻遗传改良中主要存在着两大难题,即超高产育种和杂种优势固定。在同源多倍体水平进行深入探索将有望寻找到解决这两大难题的技术思路和研究途径。深入研究同源多倍体水稻雌雄配子或雌雄配子体的发育特征、受精作用及其胚胎发育特征将有可能在短期内从根本上阐明同源多倍体水稻结实率低和籽粒充实度差的机制,进而获得结实和充实度均正常的同源多倍体水稻品系,这将有助于在染色体组多倍化水平挖掘水稻的产量潜力。对同源多倍体水稻的胚自发现象和早世代稳定现象进行深入研究,将为水稻无融合生殖研究寻找到新的突破口,这一方面可以在现有的稻种资源中进一步寻找潜在的无融合生殖基因,另一方面还可以通过物种间远缘杂交将异源物种中的无融合生殖基因转移到稻种资源中。
从染色体组的亲缘关系来看,多倍体水稻包括同源多倍体水稻、异源多倍体水稻和同源异源多倍体水稻等多种类型。从遗传组成的复杂性、研究难度和实际应用的角度来看,近期研究者首先应该对同源多倍体水稻的特征特性进行深入研究,随后再对异源多倍体水稻和同源异源多倍体水稻的特征特性进行深入研究。根据现有的资料,在研究同源多倍体水稻的过程中首先应该立足于对同源三倍体和同源四倍体水稻的特征特性及其潜在价值进行深入研究,接着再研究其他类型的同源多倍体水稻。
3 从物种间远缘杂交的生物学效应看染色体组多倍化的潜在价值
稻属植物是禾本科植物中非常重要的植物种群,它们广泛地分布于全球的热带地区和亚热带地区。关于稻属植物的生物学研究从20世纪初就已经开始,其中对其物种的类型和地理分布研究得比较清楚,对其潜在价值的挖掘主要局限在对普通栽培稻(Oryza Sativa)的遗传改良及其杂种优势利用,对非洲栽培稻(0.glaberrima)的研究深度还相当有限。由于稻属植物种类的多样性、生态条件的差异性、人类认识水平的局限性和学科发展的不平衡性,导致人类对稻属植物的研究仍然停留在一定的时代水平,这给后人留下了许多值得进一步探索和研究的难题。
3.1关于栽培稻物种间杂种在遗传上的亲和性研究
普通栽培稻和非洲栽培稻是稻属内所包含的两个栽培物种。以不同类型的普通栽培稻品种或材料与各种类型的非洲栽培稻品种或材料进行杂交的结果表明,物种间的生殖隔离并不是非常严格,在杂交数量比较大的情况下可以获得杂种第一代种子及其后代。对物种间杂种F1植株的研究结果表明,在其生殖发育过程中雌性器官发育得比较好,在其雌配子或雌配子体中有一部分发育良好,其生活力比较强而具有一定的受精能力。然而,其雄性器官发育得比较差,在其雄配子或雄配子体中大部分发育异常,其生活力和受精能力均比较弱。在普通栽培稻与非洲栽培稻杂交的大多数杂交组合中通常会表现出雌配子或雌配子体可以发生受精作用,而大多数花粉粒表现为高度不育。对于两种栽培稻远缘杂交后杂种第一代植株及其后代的特异性表现,早期的一些研究者认为,可能有一种隐藏的异源染色体结构分化中的差异性,导致了其杂种在生育特性上表现出明显的不育特征。研究者在物种间杂种植株进行减数分裂时期观察到,各种超常规的染色体异常现象(如单价体的产生、落后染色体的出现和在后期各条染色体不同步移动等),均会导致小孢子内所包含的染色体数目通常不相等,进而导致其生活力和受精能力相当弱。在研究者所考查的杂交组合中,绝大多数杂种第一代植株在减数分裂时花粉母细胞的染色体联会和四分体都表现正常,由此表明双亲的染色体具有一定的同源性,而其花粉粒败育发生在减数分裂后期。Sano(1986)利用携带不育基因的等基因系材料进行研究的结果表明,物种间杂种植株表现出高度不育性有其遗传学根源,其遗传特点涉及水稻的第一连锁群,即在该连锁群的一个特异性基因位点(Sa)上表现出“孢子体—配子体互作模式”。遗传学的检测结果证实,Sa基因与wx基因(糯性胚乳基因)在遗传上紧密连锁在一起;雄性不育基因S1和S2也位于第一连锁群,它们与C基因(稃尖色泽基因)紧密连锁;当雄性不育基因S1和S2处于隐性结合时则导致孢子体发生不育,材料的遗传背景对S1基因的表达有明显的影响。另外一类雄性不育基因(S3基因)并不会影响大孢子的正常发育,其作用主要是导致花粉粒致死。除此之外,Sano在对S1基因进行提纯的过程中偶然分离到S4雄性不育基因,它所导致的雄性不育特性属于孢子体不育类型。S4雄性不育基因与糯性胚乳基因wx分别位于不同的连锁群,因而不可能由S1基因通过基因突变产生S4基因。由此推测,S4基因的产生很可能是由特异性的转座因子所导致的结果。大量的研究结果表明,在栽培稻物种间的远缘杂种中,核质互作型雄性不育材料的出现是一个非常普遍的生物学现象,这为稻属植物雄性不育种质资源的收集和筛选提供了丰富的物质基础。
随着国内外水稻遗传育种水平的不断提高,其研究难度越来越大,迫切需要对现有的水稻种质资源进行有效的遗传改良。根据稻属分类学的研究结果,栽培稻包括两个物种,即亚洲栽培稻(普通栽培稻,Oryza sativa L.,2n=2x=24)和非洲栽培稻(光稃稻,O.glaberrima Steud,2n=2x=24)。水稻育种者长期以来都期望着将非洲栽培稻的有益性状引入到亚洲栽培稻中,以便对后者进行遗传性改良。在栽培稻的两个物种中,亚洲栽培稻的应用范围比较宽广,而非洲栽培稻的应用范围比较狭窄。然而,非洲栽培稻在米质和抗性等方面具有一些有待于进一步挖掘的优良性状。前人的研究结果表明,由于物种间的生殖隔离都比较明显,物种间的远缘杂交都比较难,远缘杂交的效率都比较低。根据我们过去对同源四倍体水稻的研究结果以及在同源四倍体水稻与狼尾草或假稻的远缘杂交中所获得的工作经验,在栽培稻的两个物种间的远缘杂交中,我们将同源四倍体水稻作为杂交母本与非洲栽培稻进行了远缘杂交配组。研究结果表明,非洲栽培稻分别与亚洲栽培稻中的两种倍性水平的水稻进行杂交均表现出一定的可交配性,生殖隔离并不是很严格。然而,不同倍性的亚洲栽培稻与非洲栽培稻杂交,其结实率表现出明显的差异。二倍体水稻与非洲栽培稻不容易杂交,其结实率比较低,结实种子的充实度比较差,而同源四倍体水稻与非洲栽培稻似乎更容易杂交,其结实率比较高,大部分结实种子的充实度比较好。
关于同源四倍体水稻与非洲栽培稻杂交的后效性尚未见到公开的文献报道。我们的研究结果表明,不同倍性的普通栽培稻与非洲栽培稻之间不存在严格的生殖隔离,通过有性杂交可以获得杂种第一代种子,但其结实率因普通栽培稻的倍性水平而异。在以同源四倍体水稻或以二倍体水稻为杂交母本,以非洲栽培稻为花粉供体所配制的32个杂交组合中,其杂种第一代群体均表现出明显的营养生长优势。从群体的生长势来看,杂种第二代群体比杂种第一代群体要弱一些;在杂种第二代群体中,在以同源四倍体水稻为杂交母本的杂交组合中所表现的分离现象比在以二倍体水稻为杂交母本的杂交组合中所表现的分离现象更加明显。在各个杂交组合的第三代群体中,从植株的株叶形态和生育期来看,株系间的差异和株系内的变异依然很明显,变异频率更宽,变异种类更多。在普通栽培稻与非洲栽培稻的杂交组合中,育性变异、生长势变异、株叶形态变异、染色体变异和结实性变异等均是比较容易发现的变异现象。根据我们的试验结果可以肯定,利用同源四倍体水稻为杂交母本与非洲栽培稻杂交更容易将后者的遗传物质引入到亚洲栽培稻,这将为普通栽培稻的遗传改良找到了一条值得进一步研究的新途径。
普通栽培稻与非洲栽培稻在遗传上存在着一定的差异性,在杂交的可交配性上并没有表现出严格的生殖隔离障碍,这为通过栽培稻物种间杂交和染色体组多倍化创造异源多倍体提供了很大的可能性。从现有的研究结果来看,在稻属植物中开展异源多倍体新物种的创造则首先应该立足于栽培稻物种间的遗传重组为重点,即以普通栽培稻和非洲栽培稻为基础,通过广泛地杂交、染色体组加倍和不断地筛选,进而获得具有实用价值的异源多倍体新材料。
3.2关于普通栽培稻与远缘物种间杂交的探索性研究
我们在研究同源四倍体水稻的生殖特性时发现,同源四倍体水稻具有比较特殊的生殖性状,其有性生殖能力比较弱,遗传可塑性比较大,在雌雄配子和雌雄配子体的发育过程中存在着明显的多态性。根据同源四倍体水稻的生殖特性,我们在以其作为杂交母本分别与狼尾草和假稻的远缘杂交中均获得了比较好的研究结果。对于这种在种间或属间远缘杂交中能比较容易地获得杂交F0种子,并且大多数F0种子的充实度能达到正常水平的特殊生殖现象,我们利用激光扫描共聚焦显微技术对其胚胎发育特征及其生殖生物学机制进行了比较深入的研究。(https://www.chuimin.cn)
自从2025年以来,一些研究者根据狼尾草(Pennisetum alopecuroides,2n=2x=18)具有生长势旺盛,穗大粒多,再生能力和宿根能力强,抗多种病虫,耐旱耐瘠等优良特性,试图通过二倍体水稻与狼尾草之间的远缘杂交方式将狼尾草的优良性状引入到水稻中,进而创造出具有特殊意义的水稻新种质。然而,在二倍体水稻与狼尾草之间的远缘杂交中生殖隔离相当严重,杂交很难成功[15]。近年来,在对多倍体水稻的研究中已经发现,其生殖性状比较特殊,有性生殖能力比较弱。在利用同源四倍体水稻为母本与狼尾草的杂交中获得了充实度达到正常水平的杂种F0种子,其杂交结实率高达2.42%~8.54%。在试验中利用激光扫描共聚焦显微技术对属间远缘杂交中的受精作用进行了研究。统计结果表明,总受精率高达6.36%~11.68%,总成胚率也高达4.11%~9.44%,这一观察结果与我们在杂交试验中所得到的结果相吻合。
关于同源四倍体水稻与假稻的远缘杂交试验主要包括两个方面的研究内容,即同源四倍体水稻与假稻的可交配性研究、同源四倍体水稻与假稻杂交结实的胚胎学机制。在杂交试验中发现,以同源四倍体水稻为杂交母本与假稻杂交则表现出一定的可交配性,由此获得了饱满的实粒种子和不饱满的结实种子,其结实率的变幅为2.29%~15.68%。胚胎学研究结果表明,二倍体水稻紫血稻(2)不能与假稻进行有性杂交的生殖障碍首先表现在假稻的花粉粒在紫血稻(2)的柱头上不能萌发。在同源四倍体水稻紫血稻(4)与假稻进行杂交的组合中所得到的观察结果则比较复杂。在授粉后30min固定的266枚颖花中,每个颖花的柱头上都沾满着假稻的花粉粒,其中,在21枚颖花(占7.89%)的柱头上发现了正在萌发的花粉粒,其花粉管正伸入柱头内,但尚未伸入到花柱内。在授粉后1.0h固定的306个颖花中,有28个颖花(占9.15%)的花柱内发现了正在伸长的花粉管。在授粉后6.0h固定的269枚颖花中,在48个颖花(占17.84%)的花柱内发现了花粉管的痕迹,在19个颖花(占7.06%)的胚囊内的珠孔端发现了花粉管进入胚囊时所留下的痕迹。这些观察结果说明,紫血稻(4)与假稻的生殖隔离不是很严格,后者的花粉粒在前者的柱头上可以萌发,花粉管可以在花柱内伸长,直至进入胚囊之内。假稻的花粉管进入胚囊后,2个精子被释放到卵细胞外的合点端。在紫血稻(4)与假稻的杂交中存在着精细胞与卵细胞的单受精现象、精细胞与次生极核的单受精现象和正常的双受精现象。统计结果表明,总受精率高达22.89%,总成胚率也高达20.82%。
同源四倍体水稻在生殖发育上的特异性值得引起关注,特别是在以栽培稻为基础的物种间远缘杂交中同源四倍体水稻是比较好的桥梁。同源四倍体水稻具有很强的遗传可塑性和很弱的遗传保守性,利用其作为水稻远缘杂交的桥梁,从野生物种中不断地引进有益的基因,由此有可能创造出具有强再生能力、强宿根性和无融合生殖特性的新种质,这将有助于杂交水稻的多代利用和固定水稻的杂种优势。以稻属内现有的种质资源为基础,利用现代生物技术创造出具有强大的种间杂种优势和实用价值的异源多倍体水稻,这不但可以促进物种升级和创造出新物种,还可能在更高层次上利用和固定其强大的杂种优势。以同源四倍体水稻为杂交母本,有望完成远缘物种间的遗传交流,进而将异源物种的有益基因引入到栽培稻中。
4 从稻属植物染色体组多倍化的研究难度看多倍体水稻的研究策略
通过现代生物技术进一步挖掘水稻的增产潜力已经成为水稻育种中的研究热点。根据杂交水稻育种的战略设想,亚种间杂种优势比品种间杂种优势更强,物种间远缘杂种优势比亚种间杂种优势更强,利用物种间远缘杂种优势是杂交水稻超高产育种的探索方向之一[16]。研究多倍体水稻所具有的一些特征特性将有助于深入开展水稻超高产育种的探索性研究,从而不断开创水稻超高产育种的新局面,在更高层次上利用水稻的杂种优势。从目前二倍体水稻的超高产育种现状来看,在二倍体水平上的增产潜力很有限,而在多倍体水平上的实用价值还有待于进一步探索。可以预言,对多倍体水稻的潜在价值还有待于进一步探索和挖掘,一旦取得突破性进展将会在水稻育种和水稻生产上掀起又一次革命性浪潮。然而,从植物物种的进化与其染色体组的关系来看,染色体组的多倍化是新物种形成的主要途径之一,而人工诱变方法的不断成熟促进了新物种形成的速度。鲍文奎等研究者一直坚信,在多倍体水平上探索稻属植物的潜在价值将会使其研究水平上升到新的高度。我们将二倍体水稻诱导为多倍体水稻并对其进行筛选的研究工作实际上是在探索稻属内新物种形成的特点和方式,其研究难度相当大,但其科学价值和对稻属科学的促进作用还无法估量。
从水稻多倍体的研究现状来看,近期迫切需要对同源多倍体水稻的一些技术问题进行深入研究。在同源多倍体水稻的研究中迫切需要形成新的技术思路,以便寻找到新的研究途径。一些研究者认为,利用离子束生物技术对同源多倍体水稻进行遗传改良,有可能为进一步利用水稻强大的杂种优势效应和深入研究水稻无融合生殖现象寻找到新的突破口[17-19]。根据目前的研究现状,对同源多倍体水稻进行遗传改良的技术思路是,瞄准2个总体目标(挖掘水稻的产量潜力和简化水稻杂种优势利用的程序),采用3种有效技术(离子束生物技术、组织培养技术和染色体组多倍化诱导技术),确定4个研究方向(直接利用同源四倍体水稻的增产潜力、利用同源四倍体水稻的强大杂种优势效应、固定其杂种优势效应和深入研究同源四倍体水稻的生殖发育特性),达到5个预期目标(探索在多倍性水平物种形成的途径、研究多倍体水稻的生物学特征特性、寻找多倍体水稻所表现的遗传变异规律、挖掘同源三倍体和同源四倍体的潜在利用价值、建立同源多倍体水稻的新的育种技术体系)。
为了挖掘同源多倍体水稻的潜在利用价值,在今后的研究中有如下5条途径值得进一步探索。其一是探索直接利用同源四倍体水稻的可能性。染色体组的多倍化使得同源四倍体水稻的特征特性发生明显的变化,其优点是大粒型、营养生长优势、生物学产量高、优良的米质和比较强的抗逆性,而其缺点主要是不良的籽粒充实度和低结实率。从现在的研究现状来看,要解决同源四倍体水稻籽粒充实度的问题并不难,而试图在获得高结实率单株之后再获得稳定遗传的高结实群体则需要进一步探索。本人认为,在挖掘同源四倍体水稻的直接利用价值的研究过程中,可以利用离子束生物技术来解决其结实率低的问题。
其二是探索在同源四倍体水平利用水稻杂种优势的可能性。对同源四倍体水平的籼粳F1杂种进行研究的结果表明,其结实率显著地高于二倍体籼粳杂种的结实率,籽粒充实度也是如此。2025年我们在利用同源四倍性原始籼稻和原始粳稻进行杂交的试验中发现,某些杂交组合的杂种第一代的结实率可以达到75%以上,最高可以达到91.45%,并且,生物学产量上的杂种优势效应很明显。然而,由于同源四倍性原始籼稻或原始粳稻本身的结实率很低,两者之间的杂交不容易成功,很难生产出大量的杂交第一代种子满足生产上的需要。在由同源四倍性原始籼稻与原始粳稻所配制的杂种第一代中存在着强大的杂种优势效应有待于我们进一步挖掘和利用,而通过利用现代生物技术对同源四倍体水稻本身的结实率性状进行遗传改良,进而筛选出结实正常的同源四倍体水稻亲本则有助于在四倍体水平利用水稻强大的杂种优势效应。
其三是探索在同源多倍体水平固定水稻杂种优势的可能性。在水稻染色体组多倍化的研究中一些研究者发现,多倍体水稻由于其本身的遗传特性和生殖发育特点,它在固定水稻杂种优势中可能会具有特殊的价值。在水稻杂种优势的普遍性得到生产实践的证实之后,为了简化水稻杂种优势利用的程序,许多研究者试图探索固定水稻杂种优势的途径和方法。然而,固定水稻杂种优势的技术难度颇大,还有许多技术性问题有待研究。从实用性来看,固定水稻杂种优势的方法包括无性繁殖法、无融合生殖法和异源多倍体法等等。当二倍体水稻被诱导为同源四倍体水稻之后,其生殖特性会发生明显的变化;同源四倍体水稻通常表现出比较弱的有性生殖能力,这主要是由于在减数分裂过程中同源染色体配对异常,出现单价体和多价体等导致配子生活力降低的不正常现象,进而使大部分雌配子体或雄配子体失去正常功能,雌配子或雄配子的生活力明显变弱。正是由于同源四倍体水稻具有比较弱的有性生殖能力,它在水稻无融合生殖研究中具有比较特殊的价值,即可以在同源四倍体水稻中筛选具有无融合生殖特性的种质资源,也可以将同源四倍体水稻作为受体,通过现代生物技术从异源物种中导入无融合生殖基因,进而筛选出具有无融合生殖特性的新种质。由此可见,以研究同源四倍体水稻为基础,有可能为研究固定水稻杂种优势的技术路线寻找到一条新途径。
其四是探索以同源多倍体水稻为桥梁完成异源遗传物质转移和丰富水稻基因库的可能性。在水稻的远缘杂交中,同源四倍体水稻是一种很好的亲本和桥梁,以其为中介可望将远缘物种中的有益基因引入到二倍体水稻中,进而创造出具有实用价值的水稻新种质。在以同源四倍体水稻为杂交母本,以狼尾草或假稻为花粉供体的属间远缘杂交中,我们在杂交当代群体内就获得了实粒种子并对其胚胎学进行了研究。尽管如此,其中的一些技术性问题仍然需要进一步研究。
其五是探索在同源多倍体水平水稻特征特性的遗传规律。根据现有的研究资料,近期对同源多倍体水稻的研究主要以同源四倍体和同源三倍体为研究材料。在研究同源四倍体水稻时主要瞄准结实性状、生物学产量潜力、籽粒产量潜力、雄性不育、异交结实率、光合特性和品质特性等性状展开研究,以便寻找其遗传规律,进而直接利用同源四倍体水稻的产量潜力和建立同源四倍体水平水稻杂种优势利用的技术程序。在研究同源三倍体水稻时主要瞄准自交结实性状的稳定性、异交结实的可交配性、雌雄配子或雌性配子体的发育特性等性状展开研究,以便寻找其生殖发育特性在亲子代之间的遗传规律,进而不断完善水稻早世代稳定育种的技术程序和探索在多倍体水平寻找水稻无融合生殖基因或种质资源的可能性。
参考文献
[1]黄群策.被子植物的无融合生殖[M].福州:福建科学技术出版社,2000.
[2]袁隆平.杂交水稻超高产育种[J].杂交水稻,2001,16(1):1-3.
[3]中国农学会,中国水稻研究所,国家杂交水稻工程技术研究中心.21世纪世界水稻遗传育种展望[M].北京:中国农业科技出版社,1999,10.
[4]蔡得田,袁隆平,卢兴桂.二十一世纪水稻育种新战略Ⅱ利用远缘杂交和多倍体双重优势进行超级稻育种[J].作物学报,2001,27(1):110-116.
[5]黄群策,孙梅元,邓启云.多倍体水稻及其潜在价值[J].杂交水稻,2001,16(1):1-3.
[6]秦瑞珍,程志军,郭秀平.利用同源四倍体花培途径创建水稻突变体群的研究[J].作物学报,2005,31(3):392-394.
[7]黄群策,孙敬三.通过异倍性水稻间杂交获得同源三倍体植株[J].植物学报,1999,41(7):741-746.
[8]谢兆辉,吴先军.水稻早世代稳定特性的遗传[J].植物分子育种,2005,3(4):469-472.
[9]谢兆辉,吴先军.来自同一个母本的两个早世代稳定群体的分析比较[J].种子,2005,24(7):23-25.
[10]周黎军,吴先军,李仕贵.水稻早世代稳定特异性的研究[J].作物学报,2005,31(6):677-685.
[11]黄群策,孙敬三.同源四倍体水稻的生殖特性研究[J].中国农业科学,1999,32(2):14-17.
[12]黄群策,梁秋霞.低能离子注入同源四倍体水稻的生物学效应[J].激光生物学报,2003,12(5):355-359.
[13]黄群策.同源四倍体水稻与假稻杂交获得实粒种子[J].作物学报,2001,27(1):133-135.
[14]黄群策,向茂成.同源四倍体水稻与假稻杂交结籽的胚胎学机制[J].作物学报,2002,28(2):286-288.
[15]王敬驹,孙敬三.水稻×狼尾草胚胎发育的细胞学观察[J].植物学报,1981,23(2):104-109.
[16]袁隆平.杂交水稻的育种战略设想[J].杂交水稻,1987,3(1):1-3.
[17]黄群策,李玉峰.离子束生物技术在水稻育种中的应用前景[J].杂交水稻,2002,17(5):53-56.
[18]黄群策.离子束介导技术在植物遗传改良上的发展趋势[J].原子核物理评论,2007,24(1):59-63.
[19]黄群策.离子束生物技术改良同源四倍体水稻的设想[J].郑州大学学报,2003,35(4):31-36.
【注释】
(1)该文曾在《杂交水稻》 [2008,23(6):1~6]刊出
相关文章

稻属植物的进化特点及其潜在价值黄群策王书玉张书艮摘要概括了稻属植物的种群特征及进化特点。关键词稻属植物;种群特征;进化特点;染色体组多倍化;产量潜力在植物物种的自然演化进程中,稻是一种起源比较早、进化程度比较高的古老植物类群。稻属植物是禾本科中非常重要的植物种群,在该属内包含有23个物种,它们广泛分布于全球的热带地区和亚热带地区。......
2025-09-30

我国超级稻育种的技术性突破促使稻属遗传改良的水平提升到新的研究层次并给水稻生产带来了革命性的巨变。通过现代生物技术进一步挖掘稻属植物的增产潜力和提高遗传改良水平已经成为水稻育种中的研究热点[7]。其二,在水稻遗传改良的研究领域内研究范围的局限性明显地限制着这一学科在挖掘稻属植物杂种优势效应上发生根本性突破。......
2025-09-30

为此,资历框架作为激发全民终身学习的新举措开始出现。互联网的发展改变了人们的学习方式,从课堂学习发展到混合式学习和在线学习,终身学习的理念转向强调优质的、灵活的、无所不在的学习,终身学习的重心转向人们的学习成效,从而促进了资历框架的兴起。......
2025-09-30

泰山的神圣地位之所以在中国古代思想界得到确认,首因应推古老五行学说的诞生及盛行。泰山在五岳中位居东方,为东岳。尽管这一理论属于牵强附会的虚妄之谈,但却得到了争霸诸侯的崇信追捧,以至使其风行天下,成为热门学问,从而进一步推高了泰山的神圣地位。其中,祭祀泰山是首位的,无形中抬高了泰山的地位。由此可见泰山政治地位的显赫。这种文化心理的形成,还与泰山地震的不断“警示”密切相关。......
2025-09-30
也就是说,实录体有自己独特的叙事方式和特有的史料价值。实录基本上属于编年体,但又于其中融入了传记,形成“编年附传”的特殊史体。北宋庆历间,王尧臣等在《崇文总目》中特立“实录类”,在史部的十三类中仅次于正史和编年而居于第三位。南宋陈振孙第一个勾画了实录体的基本特征——“编年附传”。......
2025-09-29

它是自然界物质生产过程中创造的价值,涉及自然界的消遣价值、美学价值、生命价值、科学价值以及伦理价值等。生态资源整体的使用价值表现为生态系统的使用价值,如调解气候、繁衍物种、美化净化等。生态价值是通过生态服务功能体现出来的对人类直接或间接的作用。所以,任何地区、任何国家不能忽视生态价值的系统性而孤立地发展本地或本国的经济。......
2025-09-30
例如《道情》第241小节,两组全音阶片段性的交替进行,不但没有凸显印象主义音乐的色彩,反而给多重调性的音乐环境带来了个性化的色彩对比。谱例1-6:《万年欢》451—453小节(三)全、半音阶相结合有时出现前后衔接的全音阶、半音阶交替运用,例如《道情》第344小节,主奏双簧管的三十二分音符上行全音阶在第344小节第三拍九连音的后半部分转为了半音阶的结构,构成了不同色彩的瞬间对比,刚柔并济地将音乐推至力度的g2音。......
2025-09-30
因此,纯粹主观的、完全不反映任何客观内容的评价及其标准是没有的,有的只是在反映客观内容的深刻性、全面性和准确性上的差别。实际上,人类的大多数评价标准正是这样发展的。......
2025-09-30
相关推荐